Il glicogeno è un polimero ramificato di unità di D-glucosio con funzione di deposito del monosaccaride, quindi di energia.[1]
Essendo sintetizzato senza ricorrere a uno stampo, a differenza di proteine e acidi nucleici, si presenta come una popolazione di molecole con struttura e dimensioni differenti.[2][3]
Grazie alla sua struttura ramificata, il glicogeno è una molecola compatta e solubile che esercita una debole pressione osmotica e consente un rapido rilascio del glucosio immagazzinato quando necessario.[4]
Al polimero di glucosio si associano proteine essenziali al suo metabolismo, come la glicogenina (EC 2.4.1.186), la glicogeno sintasi (EC 2.4.1.11), la glicogeno fosforilasi (EC 2.4.1.1), l’enzima deramificante (EC 2.4.1.25 e EC 3.2.1.33), e altre che ne regolano l’ancoraggio con citoscheletro e membrane. Questi aggregati polisaccaride–proteine, detti granuli β, sono presenti nel citosol di batteri, Archea, Funghi e cellule animali.
Inoltre, legati covalentemente allo scheletro polisaccaridico si trovano gruppi fosfato.[5]
Proteine e gruppi fosfato sono coinvolti nella regolazione del suo metabolismo che, come il metabolismo generale, si compone di due fasi: una fase di sintesi e una di degradazione. Nella prima, reazioni catalizzate da specifici enzimi portano alla sintesi del glicogeno, mentre nella seconda, reazioni catalizzate da specifici enzimi portano alla sua degradazione o glicogenolisi.[6]
Negli animali il glicogeno è presente in quasi tutte le cellule e, nei mammiferi, è abbondante nel fegato e muscolo scheletrico. Nel fegato diversi granuli β si associano a formare strutture superiori dette granuli α.[3]
Il glicogeno è presente anche nei lisosomi.[7]
Nell’uomo rappresenta meno dell’1% delle riserve di energia, ma è essenziale per il mantenimento dell’omeostasi glicemica.[8]
È assente nelle piante, dove l’amido costituisce la forma di riserva del glucosio. La polimerizzazione del glucosio rappresenta quindi un meccanismo universale per immagazzinare energia.[9]
Per l’uomo, il glicogeno non ha praticamente nessuna importanza nutrizionale.[10]
Indice
- Cenni storici
- Struttura chimica e molecolare del glicogeno
- Granuli β
- Granuli α
- Metabolismo del glicogeno: la fase di sintesi
- Metabolismo del glicogeno: la fase degradativa
- Regolazione coordinata del metabolismo del glicogeno
- Dove si trova il glicogeno nell’uomo
- Importanza del glicogeno per l’uomo
- Glicogeno e lavoro muscolare
- Resa energetica in condizioni anaerobiche
- Resa energetica in condizioni aerobiche
- Bibliografia
Cenni storici
Il glicogeno venne scoperto nel 1857 dal fisiologo francese Claude Bernard, considerato il fondatore della medicina sperimentale.[11] Nella seconda metà del ‘900 gli studi sul metabolismo del glicogeno hanno portato a grandi progressi della biochimica, quali scoperte come:
- la fosforilazione reversibile delle proteine;
- le protein chinasi e protein fosfatasi;
- il ruolo dei ligandi nel controllo dell’attività degli enzimi allosterici;
- l’effetto dell’insulina sull’attività degli enzimi intracellulari.[9]
Il tutto ha portato a quattro premi Nobel, di cui tre per la Fisiologia o Medicina, nell’ordine a Carl Cori e Gerty Theresa Cori, nata Radnitz, nel 1947, a Earl Sutherland Jr. nel 1971, e a Edwin Krebs ed Edmond Fischer nel 1992, e uno per la Chimica a Louis Leloir nel 1970.[12]
Struttura chimica e molecolare del glicogeno
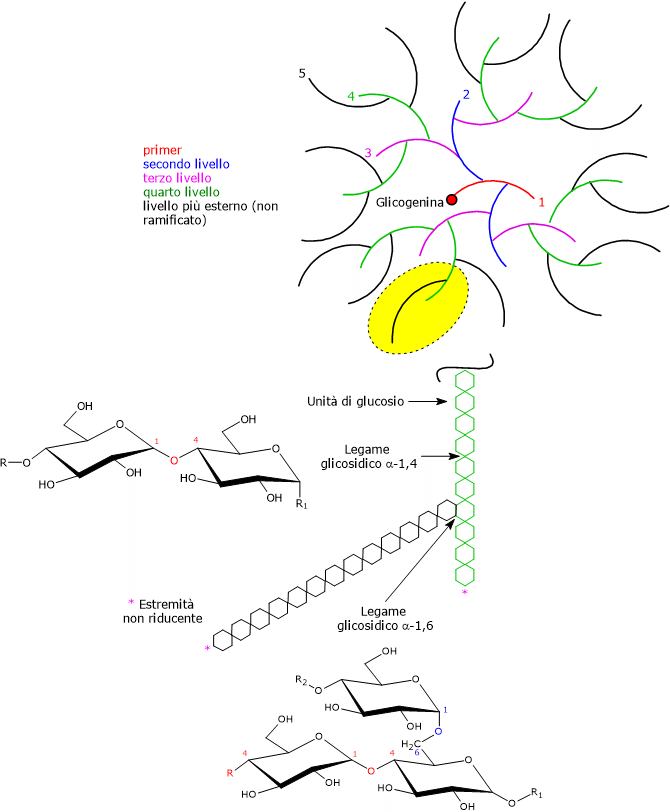
La molecola di glicogeno è un polimero ramificato di unità di D-glucosio presenti in forma piranosica, ossia anelli essenzialmente rigidi a sei atomi, cinque di carbonio e uno di ossigeno, con conformazione a sedia.
Alla catena polisaccaridica si legano covalentemente la glicogenina e gruppi fosfato.[1]
La maggior parte delle unità di glucosio sono legate attraverso legami glicosidici α-1,4, nei quali ciascuna molecola è legata alla successiva da un legame covalente tra il C-1 di una unità e il gruppo idrossilico sul C-4 dell’unità successiva, con un atomo di ossigeno che fa da ponte tra due atomi di carbonio.
I punti di ramificazione originano da legami glicosidici α-1,6 presenti all’incirca ogni 8–12 residui, di nuovo con un atomo di ossigeno a fare da ponte tra due atomi di carbonio, in questo caso C-1 e C-6. Nei mammiferi le ramificazioni hanno una lunghezza media di circa 13 residui. Poiché ogni ramificazione termina con un residuo non riducente, nella molecola saranno presenti n +1 estremità non riducenti, dove n è il numero di catene, ma solo una estremità riducente a cui è legata la glicogenina.[13]
Nota: nei disaccaridi, oligosaccaridi e polisaccaridi, l’estremità non riducente è l’estremità priva di un atomo di carbonio anomerico libero.
 Contenendo lo stesso tipo di legami glicosidici, la struttura primaria del glicogeno ricorda quella dell’amilopectina, che con l’amilosio è uno dei due polimeri di D-glucosio che formano l’amido. Tuttavia, nel glicogeno le ramificazioni sono più frequenti rispetto all’amilopectina, dove ricorrono ogni 25–30 residui, e più piccole.[14]
Contenendo lo stesso tipo di legami glicosidici, la struttura primaria del glicogeno ricorda quella dell’amilopectina, che con l’amilosio è uno dei due polimeri di D-glucosio che formano l’amido. Tuttavia, nel glicogeno le ramificazioni sono più frequenti rispetto all’amilopectina, dove ricorrono ogni 25–30 residui, e più piccole.[14]
Eterogeneità di struttura e dimensioni delle particelle di glicogeno
A differenza delle proteine e degli acidi nucleici, i polisaccaridi sono sintetizzati senza ricorrere a uno stampo, derivando dall’addizione di monosaccaridi o polisaccaridi alla struttura nascente. In aggiunta, poiché i punti di ramificazione sono disposti senza una precisa localizzazione, molecole aventi la stessa massa non necessariamente avranno la stessa struttura. Ne consegue che, all’interno di un tipo di molecole, sono presenti strutture chimiche diverse.[3]
Inoltre, molecole di glicogeno, isolate da differenti organismi, si presentano anche come una popolazione di differenti dimensioni. Risulta quindi che il meglio che si possa fare per descriverne la chimica sia definirne la distribuzione delle masse molecolari e la frequenza e lunghezza media delle ramificazioni.[2]
Infine, va sottolineato che il glicogeno non è un’entità statica, ma le sue dimensioni variano nel corso della sua esistenza.[5]
Nota: poiché il glucosio è una molecola con centri centri chirali, esiste come coppia di enantiomeri, indicati secondo la convenzione di Fischer come D-glucosio, il più diffuso in natura e monosaccaride costituente il glicogeno e l’amido, ed L-glucosio.[15]
Fattori stabilizzanti la struttura 3D
Il ripiegamento in strutture tridimensionali di macromolecole quali proteine, acidi nucleici e polisaccaridi è governato dagli stessi principi: le unità costituenti, ossia amminoacidi, nucleotidi e monosaccaridi, dotate di una struttura più o meno rigida, sono unite da legami covalenti a formare polimeri unidimensionali che spontaneamente si ripiegano a dare strutture tridimensionali stabilizzate da interazioni non covalenti quali:
- legami idrogeno;
- interazioni di van der Waals;
- interazioni idrofobiche;
- legami ionici, quando presenti unità dotate di carica.[13]
Queste interazioni possono stabilirsi all’interno di macromolecole o tra macromolecole, come nel caso delle strutture sopramolecolari come le fibre di cellulosa o i complessi multienzimatici.[16]
Dato che l’anello piranosico del glucosio è una struttura essenzialmente rigida, la conformazione tridimensionale degli oligosaccaridi e polisaccaridi da esso costituiti deriva dalla rotazione attorno ai due legami C–O del legame glicosidico, rotazione che descrive due angoli indicati come Φ (phi) e Ψ (psi). In linea teorica tale rotazione è libera, dunque gli angoli potrebbero assumere qualsiasi valore compreso tra −180° e +180°. Tuttavia questo non accade a causa dell’ingombro sterico dei sostituenti che impedisce la libera rotazione del legame C–O. Ne consegue che alcune conformazioni saranno molto più stabili di altre, determinando così la struttura tridimensionale del polimero. Sia per il glicogeno che per l’amilosio, la struttura 3D più stabile è quella di un’elica cava fortemente avvolta e stabilizzata da legami idrogeno intercatena.[9][17]
Vantaggi della struttura ramificata
La struttura altamente ramificata del glicogeno offre diversi vantaggi.
- Le estremità non riducenti presenti sul livello più esterno possono fungere da substrato per la glicogeno fosforilasi. Quindi, molte glicogeno fosforilasi possono agire simultaneamente permettendo una rapida mobilizzazione, in forma di glucoso-1-fosfato (G1P), del glucosio immagazzinato.[14]
- La struttura altamente ramificata permette al glucosio immagazzinatovi di esercitare una bassa pressione osmotica, decisamente inferiore rispetto a quella che eserciterebbe se fosse presente in forma monomerica. Se ad esempio si considera l’epatocita, vi è immagazzinata in forma di glicogeno una quantità di glucosio che in forma libera avrebbe una concentrazione di circa 0,4 M, contro una concentrazione reale di glicogeno di circa 0,01 mM. Se quindi il glucosio fosse presente in forma libera, l’osmolarità risultante sarebbe così elevata da provocare un influsso d’acqua tale da causare la lisi cellulare. Inoltre, poiché la concentrazione extracellulare del glucosio è di circa 5 mM, l’ingresso del glucosio nella cellula contro un gradiente di concentrazione tanto elevato sarebbe un processo molto dispendioso.[9]
- Le ramificazioni consentono la formazione di granuli compatti.
- Se le ramificazioni fossero assenti o comunque poche, per avere un numero di estremità non riducenti paragonabile a quelle presenti nel glicogeno, e accumulare una quantità di glucosio paragonabile, dovrebbero essere prodotti polimeri lineari molto lunghi. Questo potrebbe provocare danni alla cellula. Prove a favore di questa ipotesi arrivano da una rara malattia genetica, la malattia di Anderson, anche detta amilopectinosi o glicogenosi tipo 4, causata da mutazioni a carico del gene che codifica per l’enzima ramificante. Queste mutazioni causano una deficienza dell’attività dell’enzima e l’accumulo in diversi tessuti di glicogeno anormalmente ramificato, che ricorda l’amilopectina.[2][6]
- Le ramificazioni permettono al glicogeno di rimanere solubile, a differenza dell’amido.[5]
Il modello di Whelan
Per effetto dell’attività della glicogenina, e di seguito della glicogeno sintasi e dell’enzima ramificante, anche detto amilo-α-(1,4)→α-(1,6)-transglicosidasi (EC 2.4.1.18), la molecola del glicogeno cresce esponenzialmente in livelli concentrici attorno alla glicogenina.
Secondo il modello di Whelan della struttura del glicogeno, nella molecola si individuano due tipi di catene di glucosio:
- le catene A, non ramificate e presenti solo sulla superficie;
- le catene B, interne, con in media due ramificazioni.[18][19]
È stato calcolato che la dimensione massima del glicogeno è di 12 livelli, con un diametro di circa 42 nm, circa 55000 unità di glucosio e una massa molecolare di circa 107 Da.[20][21]
Considerando che ogni livello ha uno spessore di 3,8 nm e assumendo che la molecola di glicogeno sia sferica, dal terzo al dodicesimo livello:
- il diametro aumenta di 5,4 volte;
- il volume, che cresce in base al cubo del raggio, aumenta di 156 volte;
- il contenuto in carboidrati cresce di 45,6 volte;
- il numero di catene A in ciascun livello esterno aumenta esponenzialmente, secondo la relazione 2n −1, dove n è il numero del livello.[18]
| Livello | Diametro (nm) | Catene/Livello | Glucosio/Livello | Glucosio totale |
|---|---|---|---|---|
| 1 | — | 1 | 13 | 13 |
| 2 | 3,8 | 3 | 26 | 39 |
| 3 | 7,8 | 7 | 52 | 91 |
| 4 | 11,6 | 15 | 104 | 195 |
| 5 | 15,4 | 31 | 208 | 403 |
| 6 | 19,2 | 63 | 416 | 819 |
| 7 | 23,0 | 127 | 832 | 1651 |
| 8 | 26,8 | 255 | 1664 | 3315 |
| 9 | 30,6 | 511 | 3328 | 6643 |
| 10 | 34,4 | 1023 | 6656 | 13299 |
| 11 | 38,2 | 2047 | 13312 | 26611 |
| 12 | 42,0 | 4095 | 26624 | 53235 |
Nel muscolo scheletrico, l’analisi al microscopio elettronico delle dimensioni del glicogeno ha evidenziato la presenza di poche particelle di dimensioni massime e un diametro medio di circa 25 nm, corrispondente a sette livelli.[22]
Caratteristica del modello di Whelan
Una caratteristica importante del modello di Whelan è che il livello più esterno conterrebbe, sotto forma di catene A, circa il 50% di tutte le molecole di glucosio presenti. Ciò non significa che queste molecole siano tutte accessibili alla glicogeno fosforilasi, in quanto l’enzima si blocca a quattro residui di distanza dal punto di ramificazione. A questo punto l’enzima deramificante, la cui attività è più lenta rispetto a quella della glicogeno fosforilasi, rimuove la ramificazione rimasta permettendo alla glicogenolisi di procedere.[2][3]
Perché non è possibile il tredicesimo livello
Il tredicesimo livello sembra non essere possibile a causa dell’ingombro sterico dovuto all’elevata densità di unità di glucosio che si verrebbe a trovare sulla superficie della molecola. La densità sarebbe cioè tale da far si che non sia disponibile uno spazio sufficiente per l’interazione tra la regione catalitica degli enzimi del metabolismo del glicogeno, inclusa la glicogeno sintasi, e le catene nascenti.[2][23]
Inoltre, attraverso analisi matematiche, è stato suggerito che i valori della lunghezza delle ramificazioni, quindi circa 13 residui glucidici nei mammiferi, della frequenze medie delle ramificazione per livello, circa 2, e del numero massimo di livelli, 12, siano quelli ottimali per ottenere la mobilizzazione della quantità massima di molecole di glucosio nel più breve tempo possibile.[24]
Glicogenina
La struttura della molecola del glicogeno include la glicogenina, una proteina disposta al centro, la quale è covalentemente legata alla catena polisaccaridica. La glicogenina da inizio alla sintesi del glicogeno via autoglicosilazione, catalizzando l’addizione di 7–11 unità di glucosio a uno specifico residuo di tirosina. L’oligosaccaride neoformato funge quindi da substrato per la glicogeno sintasi. In aggiunta, la glicogenina, legandosi ai filamenti di actina, ancora il polisaccaride nascente al citoscheletro.[14][25]
Gruppi fosfato
Oltre alla glicogenina, la molecola del glicogeno lega covalentemente anche gruppi fosfato.
Per molti anni la presenza dei gruppi fosfato fu considerata una contaminazione, tanto che la quantità presente era correlata al grado di purezza del preparato. Solamente nei primi anni 80 vennero riconosciuti come parte integrante del polisaccaride, dove sembra siano legati ai C-2 e C-3 in forma di monoestere, forse come risultato di reazione collaterale nel corso dell’attività della glicogeno sintasi.[26]
Molti lavori indicano che la loro presenza abbia un ruolo nel meccanismo di regolazione del metabolismo del glicogeno, in analogia con quanto accade per il metabolismo dell’amido nelle piante. Prove a sostegno di questa ipotesi includono l’identificazione della laforina, una glicogeno fosfatasi, e il fatto che mutazioni nel suo gene sono un fattore chiave nella malattia di Lafora, una forma di epilessia caratterizzata, tra l’altro, da un’eccessiva fosforilazione del glicogeno.[27]
Ma come agirebbero i gruppi fosfato? Sono state fatte diverse ipotesi, e di seguito se ne riportano due.
- È stato suggerito che i gruppi fosfato, che sono idrofilici, possano esporre regioni idrofobiche della molecola, riducendone quindi la solubilità. Le defosforilazioni operate dalla laforina, permettendo alla molecola di rimanere solubile, faciliterebbero il processo di ramificazione.[28]
- Secondo un’altra ipotesi il grado di fosforilazione sarebbe correlato all’età della molecola, e rappresenterebbe una sorta di controllo di qualità. L’aumento delle fosforilazioni, che comporta una riduzione della solubilità della molecola, agirebbe da marker metabolico che la indirizza, anziché verso la glicogenolisi, verso la degradazione lisosomiale mediante un processo detto glicofagia.[29]
Granuli β
Le singole molecole di glicogeno sono troppo piccole per essere osservate attraverso microscopia ottica. La microscopia elettronica ha invece permesso di identificare tre tipi di strutture: i granuli β, le particelle γ, e i granuli α.[2]
I granuli β sono costituiti dal polisaccaride, dalla glicogenina e dalle particelle γ, particelle ricche di proteine con un diametro di circa 3 nm. I granuli β hanno un peso molecolare di 108 Da o più, un diametro di circa 20–30 nm e una forma simile a rosette. Sono considerate una fonte di energia rapidamente disponibile.[5][14]
In condizioni fisiologiche le proteine costituiscono il 66–80% del peso del granulo. Queste proteine stabiliscono legami anche tra di loro, con le membrane e il citoscheletro, e sono coinvolte nel metabolismo del polisaccaride. Alcune di queste sono:
- la glicogeno sintasi, l’enzima deramificante e la glicogeno fosforilasi;
- varie proteine regolatorie quali:
- le fosfatasi laforina e fosfoprotein fosfatasi 1 (PP1; EC 3.1.3.17);
- le chinasi fosforilasi chinasi (EC 2.7.11.19) e AMPK (EC 2.7.11.31);
- la proteina di ancoraggio alle membrane STDB1 (anche la fosforilasi chinasi lega le membrane);
- la malina o E3-ubiquitina ligasi (EC 2.3.2.27), che si lega al glicogeno attraverso una interazione con la laforina, e TRIM7.[30]
A differenza di quanto accade con il complesso della piruvato deidrogenasi o con i ribosomi, la composizione dei granuli β non è costante, bensì assai dinamica, in quanto le proteine si associano o dissociano dal granulo a seconda delle condizioni cellulari. In aggiunta, differenze si osservano non solo tra tipi cellulari differenti ma anche all’interno di uno stesso tipo cellulare, ad esempio nelle cellule del muscolo scheletrico in base alle loro differenti localizzazioni subcellulari.[3]
Granuli α
Nel fegato, i granuli β sono organizzati a formare un’ulteriore entità strutturale, i granuli α.
Si tratta di strutture formate da diversi granuli β, con un diametro fino a 300 nm e un peso molecolare di circa 108 kDa, e che, alla microscopia elettronica, appaiono con una forma simile ai broccoli.
I granuli α sono ritenuti una fonte di energia più lenta rispetto ai granuli β.[31]
Attualmente non è ancora chiaro il meccanismo alla base della loro formazione, anche se sembra che i granuli β siano legati attraverso uno scheletro proteico ricco in ponti disolfuro.[3]
Metabolismo del glicogeno: la fase di sintesi
Il glicogeno viene accumulato nella cellula quando l’apporto di nutrienti è adeguato. La sua sintesi avviene nel citosol, sembra essere associata ai filamenti di actina, e può avvenire a partire dal glucosio derivante dai carboidrati assunti con l’alimentazione o dal glucosio derivante da precursori non glucidici, quali acido lattico e alanina.[3][32]
Il lattato, prodotto dalle cellule muscolari scheletriche in condizioni di scarsa disponibilità di ossigeno, dai globuli rossi, che dipendono interamente dalla glicolisi anaerobica per la produzione di ATP, o derivante da altri tessuti, viene metabolizzato per la maggior parte a livello epatico per la produzione di glucosio attraverso la gluconeogenesi. Il glucosio prodotto diffonde dagli epatociti nel circolo ematico ed è trasportato ai tessuti periferici, tra cui anche il muscolo scheletrico dove può essere convertito a lattato, chiudendo così un ciclo noto come ciclo di Cori.[33]
L’alanina, un amminoacido non essenziale, può derivare da processi di transaminazione in cui il piruvato di origine glicolitica agisce da accettore del gruppo amminico di aminoacidi utilizzati a scopi energetici. L’alanina rappresenta quindi un mezzo di trasporto dello scheletro carbonioso della base coniugata dell’acido piruvico, il piruvato, e dei gruppi amminici dai tessuti extraepatici al fegato dove, lo scheletro carbonioso verrà utilizzato per produrre glucosio attraverso la gluconeogenesi e il gruppo amminico convertito in urea attraverso il ciclo dell’urea. Il glucosio diffonde quindi nel circolo sanguigno e raggiunge i tessuti extraepatici dove può essere riconvertito in piruvato chiudendo il ciclo conosciuto come ciclo glucosio-alanina.[34]
Il glucosio entra nelle cellule utilizzando specifici trasportatori proteici transmembrana chiamati GLUT, dall’inglese glucose transporter. Di questi GLUT4 prevale in tessuti insulino-dipendenti o insulino-sensibili, come il muscolo scheletrico e cardiaco e il tessuto adiposo, ed è esso stesso insulino-sensibile.[13]
Ingresso e fosforilazione del glucosio
Una volta nella cellula, il glucosio viene fosforilato a glucosio-6-fosfato (G6P). Nel fegato e nelle cellule β del pancreas questa fosforilazione è catalizzata dalla glucochinasi o esochinasi IV (EC 2.7.1.1) e dalle altre esochinasi negli altri tipi cellulari.[35]
Destino del glucosio-6-fosfato
Il destino del glucosio-6-fosfato dipende dallo stato metabolico della cellula. Può entrare nella via glicolitica per produrre energia e/o precursori per altre vie metaboliche. In alternativa può subire una isomerizzazione a glucosio-1-fosfato, un esempio di isomeria di posizione, nella reazione reversibile catalizzata dalla fosfoglucomutasi (EC 5.4.2.2). G1P può essere utilizzato per la sintesi del glicogeno, o può entrare nella via del pentoso fosfato se la cellula necessita di NADPH per le biosintesi riduttive, come quelle degli acidi grassi o del colesterolo, o se necessita di ribosio-5-fosfato per la sintesi dei nucleotidi.[9][36]
Sintesi del glicogeno
Quando prevale la sintesi del glicogeno, il glucosio-1-fosfato viene attivato a UDP-glucosio nella reazione catalizzata dalla UDP-glucosio pirofosforilasi (EC 2.7.7.9), a spese di una molecola di UTP. L’UDP-glucosio è il donatore di glucosio. Inizialmente, la glicogenina catalizza l’aggiunta di glucosio al gruppo idrossilico di Tyr194, dopodiché l’enzima aggiunge altri 6–10 residui di glucosio a formare una catena oligosaccaridica di 7–11 unità.[37] La catena oligosaccaridica agisce quindi da substrato per la glicogeno sintasi che, in combinazione con l’enzima ramificante, porta alla sintesi del glicogeno.[8]
Metabolismo del glicogeno: la fase degradativa
A differenza della sintesi del glicogeno, la fase catabolica o degradativa avviene sia nel citosol che nei lisosomi, ma con l’intervento di differenti vie metaboliche. La glicogenolisi è associata al reticolo endoplasmatico e al reticolo sarcoplasmatico, avendo come fine la liberazione del glucosio a scopi energetici.[38][39]
Le reazioni catalizzate dalla glicogeno fosforilasi, dalla α-(1,4)-glucan-6-glicosiltransferasi e dall’amilo-α-(1,6)-glucosidasi o enzima deramificante portano alla liberazione del glucosio in forma di glucosio-1-fosfato, per il 90% circa, e di glucosio libero per il restante 10%.[14]
Nella cellula muscolare scheletrica l’attività della esochinasi è così intensa che tutto il glucosio libero è immediatamente fosforilato a glucosio-6-fosfato, e dunque bloccato nella cellula stessa. Negli epatociti, nelle cellule della corteccia renale e negli enterociti, essendo presente la via gluconeogenetica, l’enzima glucosio-6-fosfatasi (EC 3.1.3.9) catalizza la defosforilazione del G6P, derivante dalla isomerizzazione del G1P, a glucosio, che può lasciare la cellula e contribuire alla regolazione della glicemia.[40]
Degradazione lisosomiale
Sebbene il glicogeno sia sintetizzato nel citosol, si ritrovato anche nei lisosomi. Il glicogeno lisosomiale sembra derivare da meccanismi di autofagia, e interessa circa il 10% del contenuto totale del glicogeno epatico e circa il 5% di quello muscolare.[41]
Secondo una ipotesi, il grado di fosforilazione del glicogeno sarebbe uno dei fattori chiave nella regolazione del suo metabolismo lisosomiale. Il grado di fosforilazione correlerebbe con l’età del granulo, rappresentando una sorta di controllo di qualità: un aumento delle fosforilazioni, con la conseguente riduzione della solubilità del granulo, costituirebbe un marker in grado di indirizzare il metabolismo del glicogeno verso la fase di degradazione lisosomiale, un processo noto anche come glicofagia.[5][42]
Nei lisosomi, la degradazione del polisaccaride è catalizzata dall’enzima α-(1,4)-glucosidasi acida (EC 3.2.1.20). L’idrolisi porta alla liberazione di D-glucosio. Poiché l’enzima idrolizza di preferenza i legami glicosidici α-(1,4), non è ancora chiaro come vengano idrolizzati i legami glicosidici α-(1,6).[43]
L’importanza del catabolismo lisosomiale del glicogeno è sottolineata dalla malattia di Pompe o glicogenosi di tipo II, causata da una mutazione del gene per la α-glucosidasi acida. La carenza di α-glucosidasi acida funzionante causa un accumulo di glicogeno nei lisosomi e in strutture vescicolari. Nella sua forma più grave, la malattia di Pompe risulta fatale entro il primo anno di vita.[7]
Regolazione coordinata del metabolismo del glicogeno
La sintesi e la degradazione del glicogeno sono processi esoergonici per cui, se operassero simultaneamente, si avrebbe uno spreco di energia. Nella cellula le due vie metaboliche sono soggette a uno stretto controllo e sono reciprocamente regolate in modo che quando una via procede l’altra rallenta. Nel corso dell’evoluzione, al pari di quanto accade per la regolazione coordinata della glicolisi e della gluconeogenesi, ciò è stato ottenuto attraverso la selezione di enzimi differenti per catalizzare i passaggi chiave delle due vie.[13]
Gli enzimi chiave in questione sono la glicogeno fosforilasi e la glicogeno sintasi, la cui attività viene regolata per mezzo di:
- modificazioni allosteriche, che avvengono in un arco temporale di millisecondi, sono istantaneamente reversibili, e chiamano in causa quali effettori allosterici gli ioni calcio, il glucosio, e metaboliti che segnalano lo stato energetico della cellula, ossia l’ATP, l’AMP e il G6P;
- modificazioni covalenti, ossia, fosforilazioni e defosforilazioni di specifiche proteine bersaglio quali, oltre ai due enzimi suddetti, la fosforilasi chinasi, fosfoprotein fosfatasi 1, e la glicogeno sintasi chinasi 3 (EC 2.7.11.26).
La regolazione covalente avviene in un arco temporale di secondi, ed è conseguente al legame di ormoni, di cui i più importanti sono l’ insulina, il glucagone e l’adrenalina o epinefrina, ai rispettivi recettori presenti sulla membrana plasmatica.[14]
Nella regolazione coordinata del metabolismo del glicogeno i meccanismi allosterici e covalenti si sovrappongono.[15]
Regolazione covalente
Di seguito sono riassunte le modificazioni metaboliche indotte dal legame di insulina, glucagone e adrenalina ai rispettivi recettori presenti sulla membrana plasmatica di epatociti e cellule muscolari scheletriche.
Insulina
L’insulina viene rilasciata dalle cellule β pancreatiche in risposta all’aumento della glicemia conseguente, ad esempio, a un pasto ricco di carboidrati, e ha un’azione anabolica. Il suo legame ai recettori presenti sulla membrana plasmatica delle cellule del fegato e del muscolo scheletrico innesca una cascata di reazioni che determinano la defosforilazione della:
- glicogeno fosforilasi, che risulta inibita;
- glicogeno sintasi, che risulta attivata.
In aggiunta, l’insulina recluta sulla membrana plasmatica delle cellule del muscolo scheletrico il trasportatore GLUT4. In questo modo viene stimolata la sintesi del glicogeno e inibita la glicogenolisi. Ciò concorre ad abbassare la glicemia.[44]
Glucagone
Il glucagone viene secreto dalle cellule α pancreatiche in risposta a una riduzione della glicemia, e ha un’azione catabolica. Il legame dell’ormone ai recettori presenti sulla membrana plasmatica degli epatociti innesca una cascata di reazioni che portano alla fosforilazione della:
- glicogeno fosforilasi, che risulta attivata;
- glicogeno sintasi, che risulta inibita.
In questo modo viene attivata la glicogenolisi e inibita la sintesi del glicogeno. Ciò concorre ad aumentare la glicemia.[9]
Adrenalina
L’adrenalina viene secreta dalle ghiandole surrenali in risposta a segnali provenienti dal sistema nervoso simpatico innescati da situazioni che provocano paura o stress, ma anche a seguito di un esercizio fisico particolarmente intenso. Al pari del glucagone ha un’azione catabolica in quanto determina la fosforilazione della:
- glicogeno fosforilasi, attivandola;
- glicogeno sintasi, inibendola.
Tuttavia, a differenza del glucagone agisce sia sul fegato che sul muscolo scheletrico, a seguito del legame ai recettori adrenergici β2 nel muscolo scheletrico, e ai recettori adrenergici β2 e α1 nel fegato.[15]
Vasopressina
La vasopressina e l’adrenalina, quando si legano ai recettori α1 adrenergici, innescano vie intracellulari che portano al rilascio di ioni calcio dal reticolo endoplasmatico, stimolando quindi stimolazione della glicogenolisi e inibizione della sintesi del glicogeno.[8]
Regolazione allosterica
Di seguito sono riassunte le modificazioni metaboliche indotte dal legame degli effettori allosterici AMP, ATP, G6P, glucosio e ioni calcio ai rispettivi siti di legame presenti sugli enzimi bersaglio nel muscolo scheletrico e nel fegato.
Muscolo scheletrico
A seguito di un aumento della concentrazione dell’AMP:
- la glicogeno fosforilasi b è attivata.
A seguito di un aumento della concentrazione dell’ATP:
- la glicogeno fosforilasi b è inibita.
In risposta a un aumento della concentrazione del glucosio-6-fosfato:
- PP1 è attivata;
- la glicogeno fosforilasi b è inibita;
- la forma fosforilata della glicogeno sintasi, ossia la glicogeno sintasi b, è attivata allostericamente, a seguito di un cambio conformazionale che favorisce la defosforilazione per opera di PP1. Questa attivazione permette alla glicogeno sintasi di agire da sensore per il glucosio-6-fosfato.
Quindi, quando le concentrazioni dell’ATP e del glucosio-6-fosfato sono basse ed è alta quella dell’AMP, la glicogeno sintasi è inibita, glicogeno fosforilasi è stimolata, e di conseguenza la glicogenolisi è stimolata mentre la sintesi del glicogeno è inibita.
Di contro, quando la concentrazione dell’ATP e del glucosio-6-fosfato sono elevate, viene stimolata la sintesi del glicogeno e inibita la glicogenolisi.[5]
Nelle cellule muscolari scheletriche l’aumento della concentrazione intracellulare di ioni calcio, rilasciato dal reticolo sarcoplasmatico, innesca la contrazione muscolare. Inoltre, ioni calcio si legano alla calmodulina, che è la subunità δ della fosforilasi chinasi muscolare. Il complesso ioni calcio-calmodulina attiva la chinasi, che fosforila la glicogeno fosforilasi e la glicogeno sintasi, attivando la prima e inibendo la seconda.[9]
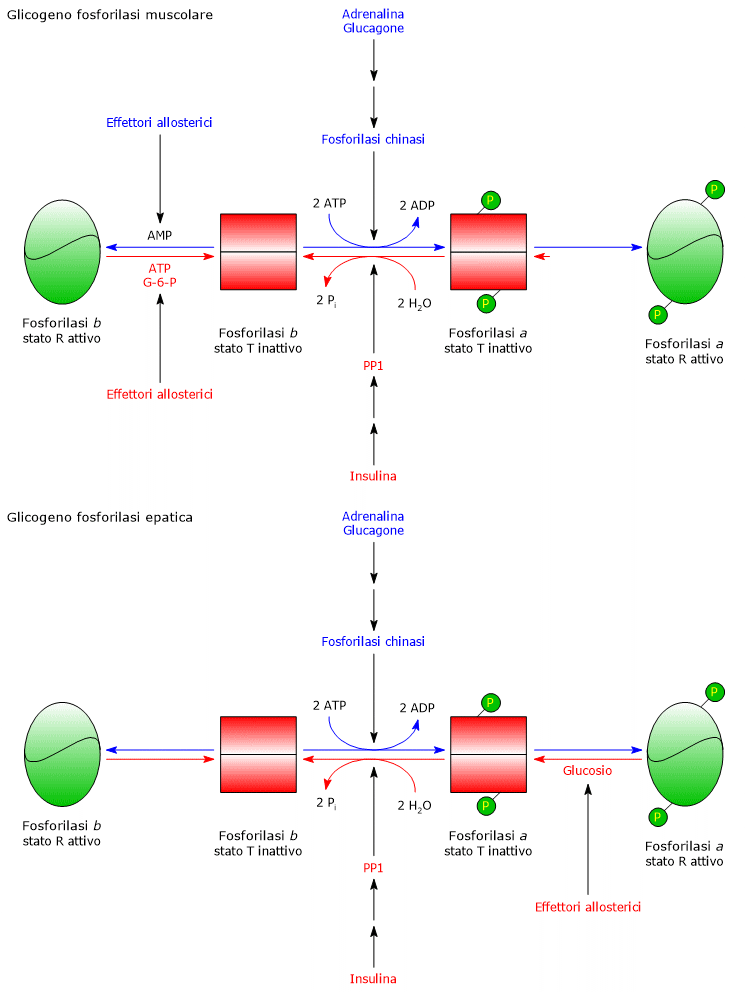
Fegato
Nel fegato la glicogeno fosforilasi b non è attivata dall’AMP, mentre la glicogeno fosforilasi a è inattivata da un aumento della glicemia.
La concentrazione del glucosio nel fegato riflette quella nel sangue: in risposta a un aumento della glicemia il glucosio si lega a uno specifico sito allosterico inibitore della glicogeno fosforilasi a e innesca una modificazione conformazionale che porta all’esposizione di specifici residui di serina fosforilati che vengono defosforilati da PP1, con conseguente inattivazione dell’enzima. Ciò permette alla glicogeno fosforilasi di agire come sensore del glucosio, e di rispondere in modo adeguato alle sue variazioni ematiche.[9]
Dove si trova il glicogeno nell’uomo
Nell’uomo il glicogeno è presente in tutte le cellule. Tuttavia i principali depositi si trovano nel fegato e nel muscolo scheletrico, dove, a seconda dello stato nutrizionale, il polisaccaride può rappresentare fino al 10% della massa del fegato e al 2% di quella del muscolo.[6] Quindi, il muscolo rispetto al fegato ha una limitata capacità di immagazzinare glicogeno. Tuttavia, poiché la sua massa è maggiore rispetto a quella del fegato, la quantità presente nel muscolo è circa il doppio di quella che si ritrova nel fegato. Ad esempio, in un soggetto adulto maschio di 70 kg non a digiuno ci sono circa 250 g di glicogeno muscolare e circa 100 g di glicogeno epatico.[45] Questi valori possono leggermente aumentare negli atleti, come nei migliori maratoneti maschi dove le riserve, tra muscolo e fegato, arrivano fino a circa 475 g, pari a circa 1900 kcal.[46]
Le quantità di glicogeno immagazzinate sono decisamente inferiori rispetto a quelle dei grassi perché questi ultimi sono una forma di deposito di energia molto più efficiente. Ciò è dovuto a diversi fattori.
- I grassi possono essere immagazzinati in forma anidra, mentre il glicogeno lega una quantità d’acqua pari a circa 2–3 volte il suo peso.
- Essendo i grassi accumulati insolubili in acqua sono osmoticamente inerti.
- Dall’ossidazione di un grammo di grasso si ottiene circa il doppio delle kilocalorie derivanti dall’ossidazione di un grammo di glicogeno: 9 vs 4.[8]
Importanza del glicogeno per l’uomo
I grassi, le proteine e il glicogeno sono riserve di energia cui l’organismo attinge in caso di necessità.
Negli animali, i grassi sono secondi solo alle proteine come riserva energetica, sebbene queste siano utilizzate solo in condizioni estreme, come durante un digiuno prolungato.
In un adulto normopeso, i grassi rappresentano circa il 21% del peso corporeo nell’uomo e il 26% nella donna. In un uomo di 70 kg, la massa grassa può soddisfare il fabbisogno energetico per circa due mesi, mentre le riserve di glicogeno bastano solo per circa un giorno. Tuttavia, il glicogeno viene comunque immagazzinato.[8]
Perché?
- A differenza del glucosio, gli acidi grassi non possono essere metabolizzati in assenza di ossigeno, e quindi non forniscono energia al muscolo durante l’attività anaerobica. Inoltre, la resa energetica del glicogeno, o meglio del glucosio da esso derivato, varia tra condizioni anaerobiche e aerobiche, e il muscolo non riesce a mobilitare gli acidi grassi con la stessa rapidità del glucosio immagazzinato.[14]
- Gli animali non possono convertire gli acidi grassi in glucosio, quindi questi non contribuiscono al mantenimento della glicemia. Sebbene nel muscolo il glucosio derivato dal glicogeno rimanga nella cellula, nel fegato, e in misura minore nel rene, grazie alla glucosio-6-fosfatasi, entra nella circolazione sistemica contribuendo all’omeostasi glicemica.[40]
- Durante la vita fetale, il glicogeno svolge un ruolo importante nelle cellule polmonari di tipo II che, intorno alla 26° settimana di gestazione iniziano ad accumularlo per la sintesi del surfattante polmonare, dove è utilizzato per la sintesi dei lipidi costituenti.[47][48]
- Il cervello, piccoli depositi di glicogeno, localizzati principalmente negli astrociti, si accumulano durante il sonno e sono mobilizzati al risveglio, suggerendo un ruolo funzionale nel cervello cosciente. Questa riserva di glucosio fornisce anche una moderata protezione contro l’ipoglicemia.[49][50]
Glicogeno e lavoro muscolare
I carboidrati, ossia il glucosio, e gli acidi grassi rappresentano le principali fonti di energia per il muscolo durante l’attività fisica, e il loro contributo relativo varia in funzione dell’intensità e durata dell’esercizio, come descritto di seguito:
- <30% VO2max: principalmente acidi grassi;
- 40-60% VO2max: acidi grassi e carboidrati in egual misura;
- 75% VO2max: principalmente carboidrati;
- >80% VO2max: circa 100% carboidrati;
Quindi, il contributo del glicogeno al totale dell’energia necessaria per sostenere il lavoro muscolare aumenta con l’aumentare dell’intensità dell’esercizio, mentre si riduce quello degli acidi grassi.
Inoltre, se non sono assunti carboidrati, la prestazione è determinata dalle riserve endogene di glicogeno muscolare ed epatico, il cui consumo relativo è differente: all’aumentare dell’intensità dell’esercizio aumenta il consumo del glicogeno muscolare, mentre il consumo del glicogeno epatico rimane più o meno costante.
Nota: il contributo relativo di acidi grassi e glicogeno come fonti di energia varia anche in funzione del livello di allenamento dell’atleta.[8][50]
Resa energetica in condizioni anaerobiche
In condizioni anaerobiche, l’ossidazione di una molecola di glucosio a lattato attraverso la glicolisi anaerobica porta alla produzione di due molecole di ATP.
Di seguito viene analizzata la resa in ATP dall’ossidazione anaerobica del glucosio rilasciato nel corso della glicogenolisi.
Glicogeno fosforilasi e ossidazione del G1P in condizioni anaerobiche
La sintesi del glicogeno dal glucosio comporta il consumo di due molecole di ATP per ogni molecola immagazzinata.
Il rilascio di una molecola di glucosio-1-fosfato per azione della glicogeno fosforilasi permette il risparmio di una delle due molecole di ATP utilizzate nella fase preparatoria della glicolisi. L’ossidazione anaerobica del glucosio-6-fosfato, prodotto dal G1P nella reazione catalizzata dalla fosfoglucomutasi, porta alla produzione di tre molecole di ATP e non due, in quanto:
- nella fase preparatoria della glicolisi viene consumata una molecola di ATP e non due, essendo bypassata la reazione catalizzata dalla esochinasi;
- quattro molecole di ATP sono prodotte nella fase di recupero energetico della glicolisi.
Il rapporto spesa-guadagno è 1/3, quindi si ha una resa energetica di circa il 66,7%.
La reazione complessiva è:
Glicogeno(n residui di glucosio) + 3 ADP + 3 Pi → Glicogeno(n−1 residui di glucosio) + 2 Lattato + 3 ATP
Considerando le due molecole di ATP spese nel corso della sintesi del glicogeno e l’ossidazione anaerobica del glucosio-1-fosfato a lattato, si ha una resa pari a una molecola di ATP per molecola di glucosio immagazzinata.
La reazione complessiva è:[51]
Glucosio + ADP + Pi → 2 Lattato + ATP
Enzima deramificante e ossidazione del glucosio in condizioni anaerobiche
Considerando il glucosio liberato per azione dell’enzima deramificante, la resa in ATP è pari a zero in quanto:
- la sintesi del glicogeno dal glucosio comporta la spesa di due molecole di ATP;
- l’enzima deramificante libera glucosio, dunque nella fase preparatoria della glicolisi saranno spese due molecole di ATP;
- quattro molecole di ATP sono prodotte nella fase di recupero energetico della glicolisi.
Se ora si considera l’ossidazione sino a lattato di tutto il glucosio provenienti dal glicogeno si ha una resa energetica pari a:
1 − [ (1/3 × 0,9) + (1 × 0,1) ] = 0,60
Ne consegue che in condizioni anaerobiche, si ha una resa energetica pari al 60%, il che indica che il glicogeno è forma efficiente di riserva di energia.[51]
Resa energetica in condizioni aerobiche
In condizioni aerobiche, l’ossidazione di una molecola di glucosio a CO2 e H2O attraverso glicolisi, complesso della piruvato deidrogenasi, ciclo di Krebs, catena di trasporto degli elettroni mitocondriale e fosforilazione ossidativa porta alla produzione di circa 30 molecole di ATP.[9]
Di seguito viene analizzata la resa in ATP dall’ossidazione aerobica del glucosio rilasciato dal glicogeno per azione della glicogeno fosforilasi e dell’enzima deramificante.
Glicogeno fosforilasi e ossidazione del G1P in condizioni aerobiche
Dall’ossidazione del glucosio-6-fosfato, prodotto dal glucosio-1-fosfato per azione della fosfoglucomutasi, a CO2 e H2O si ottengono 31 molecole di ATP, e non 30, grazie all’ATP risparmiato nella fase preparatoria della glicolisi. Dunque, il rapporto spesa-guadagno è di 1/31, corrispondente a una resa energetica di circa il 97%.
La reazione complessiva è:
Glicogeno(n residui di glucosio) + 31 ADP + 31 Pi → Glicogeno(n−1 residui di glucosio) + 31 ATP + 6 CO2 + 6 H2O
Considerando le due molecole di ATP spese per molecola di glucosio immagazzinata nel glicogeno e l’ossidazione aerobica del glucosio-1-fosfato a CO2 e H2O, si ha una resa pari a 29 molecole di ATP per unità di glucosio immagazzinata.
La reazione complessiva è:[51]
Glucosio + 29 ADP + 29 Pi → 29 ATP + 6 CO2 + 6 H2O
Enzima deramificante e ossidazione del glucosio in condizioni aerobiche
Considerando le unità di glucosio liberate dall’enzima deramificante, la resa è pari a 30 molecole di ATP in quanto la fase preparatoria della glicolisi comporta la spesa di due molecole di ATP. Quindi, il rapporto spesa-guadagno è 2/30, corrispondente a una resa energetica di circa il 93,3%.
Se ora si considera l’ossidazione a CO2 e H2O di tutte le unità di glucosio provenienti dal glicogeno, la resa energetica è pari a:
1 − [1/31 × 0,9] + [2/30 × 0,1] = 0,96
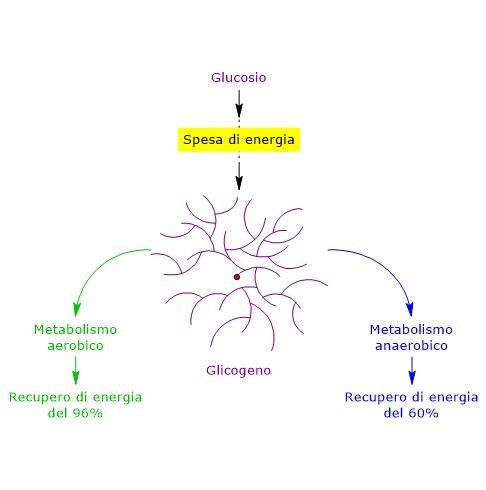
Pertanto, in condizioni aerobiche, si ha una resa energetica pari al 96%, il che indica che il glicogeno è una forma di riserva di energia estremamente efficiente, con un guadagno del 36% rispetto alle condizioni anaerobiche.[51]
Bibliografia
- ^ a b Daghlas S.A., Mohiuddin S.S. Biochemistry, Glycogen. [Updated 2023 May 1]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK539802/
- ^ a b c d e f Roach P.J., Depaoli-Roach A.A., Hurley T.D, Tagliabracci V.C. Glycogen and its metabolism: some new developments and old themes. Biochem J 2012;441:763-87. doi:10.1042/BJ20111416
- ^ a b c d e f g Neoh G.K.S., Tan X., Chen S., Roura E., Dong X., Gilbert R.G. Glycogen metabolism and structure: a review. Carbohydr Polym 2024;346:122631. doi:10.1016/j.carbpol.2024.122631
- ^ Michal G., Schomburg D. Biochemical pathways. An atlas of biochemistry and molecular biology. 2nd Edition. John Wiley J. & Sons, Inc. 2012
- ^ a b c d e f Prats C., Graham T.E., and Shearer J. The dynamic life of the glycogen granule. J Biol Chem 2018;293(19):7089-98. doi:10.1074/jbc.R117.802843
- ^ a b c Adeva-Andany M.M., González-Lucán M., Donapetry-García C., Fernández-Fernández C., Ameneiros-Rodríguez E. Glycogen metabolism in humans. BBA Clin 2016;27;5:85-100. doi:10.1016/j.bbacli.2016.02.001
- ^ a b Canibano-Fraile R., Harlaar L., Dos Santos C.A., et al. Lysosomal glycogen accumulation in Pompe disease results in disturbed cytoplasmic glycogen metabolism. J Inherit Metab Dis 2023;46(1):101-115. doi:10.1002/jimd.12560
- ^ a b c d e f Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 4th Edition. St. Louis: Elsevier, 2018.
- ^ a b c d e f g h i Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 8th Edition. W.H. Freeman and Company, 2021.
- ^ Terlouw E.M.C., Picard B., Deiss V., Berri C., Hocquette J.F., Lebret B., Lefèvre F., Hamill R., Gagaoua M. Understanding the determination of meat quality using biochemical characteristics of the muscle: stress at slaughter and other missing keys. Foods 2021;10(1):84. doi:10.3390/foods10010084
- ^ Claude Bernard, the founder of modern medicine. Cells 2022;11(10):1702. doi:10.3390/cells11101702
- ^ The Nobel Foundation. Retrieved October 30, 2025. https://www.nobelprize.org/the-nobel-prize-organisation/the-nobel-foundation/
- ^ a b c d Berg J.M., Tymoczko J.L., Gregory J.G. Jr, Stryer L. Biochemistry. 9th Edition. W.H. Freeman and Company, 2019.
- ^ a b c d e f g Heilman D., Woski S., Voet D., Voet J.G., Pratt C.W. Fundamentals of biochemistry: life at the molecular level. 6th Edition. Wiley, 2023.
- ^ a b c Garrett R.H., Grisham C.M. Biochemistry. 6th Edition. Brooks/Cole, Cengage Learning, 2016.
- ^ Alberts B., Johnson A., Lewis J., Morgan D., Raff M., Roberts K., Walter P. Molecular Biology of the Cell. 7th Edition. Garland Science, Taylor & Francis Group, 2022.
- ^ Yu Y., Delbianco M. Conformational studies of oligosaccharides. Chemistry 2020;26(44):9814-9825. doi:10.1002/chem.202001370
- ^ a b Gunja-Smith Z., Marshall J.J., Mercier C., Smith E.E., Whelan W.J. A revision of the Meyer-Bernfeld model of glycogen and amylopectin. FEBS Lett 1970:12(2);101-104. doi:10.1016/0014-5793(70)80573-7
- ^ Whelan W.J. Enzymic explorations of the structures of starch and glycogen. Biochem J 1971;122(5):609-622. doi:10.1042/bj1220609
- ^ Meléndez-Hevia E., Waddell T.G., Shelton E.D. Optimization of molecular design in the evolution of metabolism: the glycogen molecule. Biochem J 1993;295(2):477-83. doi:10.1042/bj2950477
- ^ Shearer J., Graham T.E. Novel aspects of skeletal muscle glycogen and its regulation during rest and exercise. Exerc Sport Sci Rev 2004;32(3):120-6. doi:10.1097/00003677-200407000-00008
- ^ Marchand I., Chorneyko K., Tarnopolsky M., Hamilton S., Shearer J., Potvin J., Graham T.E. Quantification of subcellular glycogen in resting human muscle: granule size, number, and location. J Appl Physiol (1985) 2002;93(5):1598-607. doi:10.1152/japplphysiol.00585.2001
- ^ Meléndez R., Meléndez-Hevia E., Mas F., Mach J., Cascante M. Physical constraints in the synthesis of glycogen that influence its structural homogeneity: a two-dimensional approach. Biophys J 1998;75(1):106-14. doi:10.1016/S0006-3495(98)77498-3
- ^ Melendez R., Melendez-Hevia E., Cascante M. How did glycogen structure evolve to satisfy the requirement for rapid mobilization of glucose? A problem of physical constraints in structure building. J Mol Evol 1997;45(4):446-55. doi:10.1007/PL00006249
- ^ Fastman N.M., Liu Y., Ramanan V., Merritt H., et al. The structural mechanism of human glycogen synthesis by the GYS1-GYG1 complex. Cell Rep 2022;40(1):111041. doi:10.1016/j.celrep.2022.111041
- ^ Fontana J.D. The presence of phosphate in glycogen. FEBS Lett 1980;1:109(1):85-92. doi:10.1016/0014-5793(80)81317-2
- ^ Gentry M.S., Guinovart J.J., Minassian B.A., Roach P.J., Serratosa J.M. Lafora disease offers a unique window into neuronal glycogen metabolism. J Biol Chem 2018;293(19):7117-25. doi:10.1074/jbc.R117.803064
- ^ Turnbull J., Tiberia E., Pereira S., et al. Deficiency of a glycogen synthase-associated protein, Epm2aip1, causes decreased glycogen synthesis and hepatic insulin resistance. J Biol Chem 2013;288(48):34627-37. doi:10.1074/jbc.M113.483198
- ^ Roach P.J. Glycogen phosphorylation and Lafora disease. Mol Aspects Med 2015;46:78-84. doi:10.1016/j.mam.2015.08.003
- ^ Brewer M.K., Gentry M.S. Brain glycogen structure and its associated proteins: past, present and future. Adv Neurobiol 2019;23:17-81. doi:10.1007/978-3-030-27480-1_2
- ^ Sullivan M.A., Aroney S.T., Li S., Warren FJ, et al. Changes in glycogen structure over feeding cycle sheds new light on blood-glucose control. Biomacromolecules 2014;15(2):660-5. doi:10.1021/bm401714v
- ^ Baqué S., Guinovart J.J., Ferrer J.C. Glycogenin, the primer of glycogen synthesis, binds to actin. FEBS Lett 1997;417(3):355-9. doi:10.1016/s0014-5793(97)01299-4
- ^ Katz J., Tayek J.A. Gluconeogenesis and the Cori cycle in 12-, 20-, and 40-h-fasted humans. Am J Physiol 1998;275(3):E537-42. doi:10.1152/ajpendo.1998.275.3.E537
- ^ Felig P., Pozefsk T., Marlis E., Cahill G.F. Alanine: key role in gluconeogenesis. Science 1970;167(3920):1003-1004. doi:10.1126/science.167.3920.1003
- ^ Wilson J.E. Isozymes of mammalian hexokinase: structure, subcellular localization and metabolic function. J Exp Biol 2003;206(Pt 12):2049-57. doi:10.1242/jeb.00241
- ^ Horecker B.L. The pentose phosphate pathway. J Biol Chem 2002;277(50):47965-47971. doi:10.1074/jbc.X200007200
- ^ Smythe C., Cohen P. The discovery of glycogenin and the priming mechanism for glycogen biogenesis. Eur J Biochem 1991;200(3):625-31. doi:10.1111/j.1432-1033.1991.tb16225.x
- ^ Zois C.E., Harris A.L. Glycogen metabolism has a key role in the cancer microenvironment and provides new targets for cancer therapy. J Mol Med (Berl) 2016;94(2):137-54. doi:10.1007/s00109-015-1377-9
- ^ Mandl J. Glycogen-endoplasmic reticulum connection in the liver. Int J Mol Sci 2023;24(2):1074. doi:10.3390/ijms24021074
- ^ a b Rajas F., Gautier-Stein A., Mithieux G. Glucose 6-phosphate, a central hub for liver carbohydrate metabolism. Metabolites 2019;9(12):282. doi:10.3390/metabo9120282
- ^ Zirin J., Nieuwenhuis J., Perrimon N. Role of autophagy in glycogen breakdown and its relevance to chloroquine myopathy. PLoS Biol 2013;11(11):e1001708. doi:10.1371/journal.pbio.1001708
- ^ Koutsifeli P., Varma U., Daniels L.J., et al. Glycogen-autophagy: molecular machinery and cellular mechanisms of glycophagy. J Biol Chem 2022;298(7):102093. doi:10.1016/j.jbc.2022.102093
- ^ Okuyama M., Saburi W., Mori H., Kimura A. α-Glucosidases and α-1,4-glucan lyases: structures, functions, and physiological actions. Cell Mol Life Sci 2016;73(14):2727-51. doi:10.1007/s00018-016-2247-5
- ^ Rahman M.S., Hossain K.S., Das S., et al. Role of insulin in health and disease: an update. Int J Mol Sci 2021;22(12):6403. doi:10.3390/ijms22126403
- ^ Knuiman P., Hopman M.T., Mensink M. Glycogen availability and skeletal muscle adaptations with endurance and resistance exercise. Nutr Metab (Lond) 2015;12:59. doi:10.1186/s12986-015-0055-9
- ^ Beelen M., Burke L.M., Gibala M.J., van Loon J.C. Nutritional strategies to promote postexercise recovery. Int J Sport Nutr Exerc Metab 2010:20(6);515-32. doi:10.1123/ijsnem.20.6.515
- ^ Farrell P.M., Bourbon J.R. Fetal lung surfactant lipid synthesis from glycogen during organ culture. Biochim Biophys Acta 1986;878(2):159-67. doi:10.1016/0005-2760(86)90142-6
- ^ Yildiz Atar H., Baatz J.E., Ryan R.M. Molecular mechanisms of maternal diabetes effects on fetal and neonatal surfactant. Children (Basel) 2021;8(4):281. doi:10.3390/children8040281
- ^ Rosenthal M.D., Glew R.H. Medical Biochemistry − Human Metabolism in Health and Disease. John Wiley J. & Sons, Inc., 2009.
- ^ Duran J., Gruart A., López-Ramos J.C., Delgado-García J.M., Guinovart J.J. Glycogen in astrocytes and neurons: physiological and pathological aspects. Adv Neurobiol 2019;23:311-329. doi:10.1007/978-3-030-27480-1_10
- ^ a b c d Snider M.D., Croniger C.M. Devlin′s textbook of biochemistry with clinical correlations. 8th Edition. John Wiley & Sons, Inc. 2024.