I lipidi, con i carboidrati, le proteine e gli acidi nucleici, sono una delle quattro principali classi di molecole organiche presenti negli organismi viventi.
Sono una classe estremamente ampia che comprende molecole con strutture differenti, riguardo agli atomi che le costituiscono, ai tipi di legami covalenti e alla presenza o meno di strutture cicliche. Esempi includono gli acidi grassi, i trigliceridi, i fosfolipidi, i glicolipidi, gli steroli, come il colesterolo, i terpeni, come i carotenoidi, e le vitamine A, D, E e K, definite vitamine liposolubili.
I diversi lipidi hanno proprietà molto varie; ad esempio alcuni sono solubili in solventi non polari, altri sono anfipatici e parzialmente solubili in alcuni solventi polari.
Con i carboidrati e le proteine, sono uno dei tre macronutrienti dell’organismo dove svolgono diverse funzioni, come di deposito di energia, di isolante elettrico e termico, di componente delle membrane, e di regolazione delle attività cellulari agendo come secondi messaggeri, ormoni e recettori di membrana. Inoltre facilitano i processi digestivi.
La digestione dei lipidi è complicata dalla insolubilità di molti di essi nell’ambiente acquoso intestinale, e per questo richiede l’intervento, oltre che di specifici enzimi, dei sali biliari. Anche l’assorbimento dei lipidi, o più correttamente l’assorbimento dei prodotti della loro digestione, è influenzato dalla solubilità delle molecole rilasciate, al pari del loro successivo trasporto, immagazzinamento e utilizzo.
In Sintesi: Punti Chiave
- Natura chimica: molecole idrofobiche e anfipatiche, solubili in solventi non polari.
- Classificazione: suddivisi in semplici (trigliceridi), complessi (fosfolipidi) e derivati (acidi grassi e steroli).
- Funzioni: riserva energetica (9 kcal/g), struttura delle membrane, isolamento termico e segnalazione ormonale.
- Nutrienti essenziali: Include le vitamine liposolubili (A, D, E, K) e gli acidi grassi essenziali (Omega-3 e Omega-6).
- Digestione e assorbimento: necessita di sali biliari e formazione di micelle miste per l’assorbimento.
Indice
- Struttura
- Dimensioni
- Solubilità
- Classificazione dei lipidi
- Funzioni dei lipidi
- Digestione
- Assorbimento intestinale
- Bibliografia
Struttura
I lipidi più semplici sono gli acidi grassi, che sono acidi carbossilici con una catena carboniosa di lunghezza variabile. Nella maggior parte dei casi la catena carboniosa è priva di ramificazioni. Può essere priva di doppi, o raramente tripli, legami, come nel caso degli acidi grassi saturi, o contenerne uno o più, e in questo caso si parla di acidi grassi insaturi. Quando la catena idrocarburica contiene due o più doppi legami, gli acidi grassi sono detti polinsaturi.
Esempi di lipidi più complessi sono i trigliceridi, i fosfolipidi e gli steroli. I trigliceridi si ottengono quando gli acidi grassi sono esterificati alla molecola del glicerolo. Se alla molecola del glicerolo vengono esterificati, oltre agli acidi grassi, anche un acido ortofosforico si ottengono i fosfolipidi; a sua volta il fosfato può legare una molecola organica quale la serina, l’inositolo, la colina o l’etanolammina. Gli steroli differiscono dagli altri lipidi per la presenza di strutture cicliche interconnesse, cui si legano catene laterali costituite da atomi di ossigeno, carbonio e idrogeno.
Dimensioni
A differenza di molti polisaccaridi, delle proteine e degli acidi nucleici, i lipidi non sono polimeri ma molecole di dimensioni contenute, con peso molecolare che varia molto ed è compreso tra circa 100 e 1.500 Dalton (Da). Ad esempio, l’acido acetico, il più piccolo tra gli acidi grassi, ha un peso molecolare pari a 60,05 Da, l’acido lignocerico, uno degli acidi grassi a catena molto lunga ha un peso molecolare di 368,37 Da, mentre quello delle proteine può variare da 5.000−10.000 Da per le più piccole, fino ad alcuni milioni per le più grandi.
Solubilità
I lipidi sono generalmente considerati molecole scarsamente solubili o insolubili in solventi polari, come l’etanolo e l’acqua, e solubili in solventi non polari, e per molti di loro questo è vero. Tuttavia, la presenza di atomi quali l’ossigeno, e soprattutto fosforo, ha l’effetto di aumentare la solubilità della molecola, o almeno di parte di essa, nei solventi polari. Considerando ad esempio gli acidi grassi saturi a catena lineare, la loro solubilità in solventi polari è funzione della lunghezza della catena idrocarburica, che ha carattere non polare, a differenza del gruppo carbossilico che è polare. L’acido butirrico, uno degli acidi grassi a catena corta, è solubile in solventi polari, mentre dall’acido caproico in avanti, passando per l’acido caprilico, l’acido caprico e l’acido laurico, le cui catene idrocarburiche sono composte rispettivamente da 6, 8, 10, e 12 atomi, la solubilità va gradualmente diminuendo.
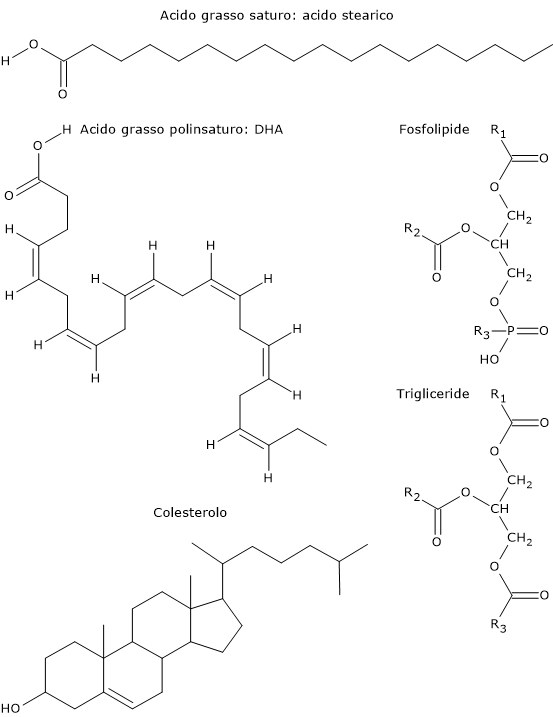
Gli acidi grassi saturi con una catena superiore ai 16−18 atomi di carbonio, quindi dall’acido palmitico e acido stearico in avanti, passando per l’acido arachidico, l’acido beenico e l’acido lignocerico, sono invece insolubili in solventi polari. Altri esempi di lipidi idrofobici sono i trigliceridi contenenti acidi grassi a catena lunga e molto lunga e gli esteri del colesterolo.
Va infine sottolineato che molti lipidi sono molecole anfipatiche, ossia dotate di una parte solubile in solventi polari e una solubile in solventi non polari. Esempi sono gli acidi grassi, i fosfolipidi, i glicolipidi e il colesterolo.
Classificazione dei lipidi
Come per la classificazione delle proteine e dei carboidrati, ci sono diversi modi per classificare i lipidi. Possono essere classificati sulla base delle loro proprietà fisiche a temperatura ambiente, quindi come solidi o liquidi, e in questo caso si parla rispettivamente di grassi e oli, o sulla base della polarità. Tuttavia la classificazione preferibile si basa sulla loro struttura chimica, considerando la quale si possono individuare tre gruppi principali: i lipidi semplici, i lipidi complessi e i lipidi derivati.
Lipidi semplici
I lipidi semplici sono formati da due tipi di unità strutturali, e includono:
- gli esteri di glicerolo e acidi grassi, come i trigliceridi, i digliceridi e i monogliceridi;
- gli esteri di colesterolo e acidi grassi;
- le cere, che sono esteri di alcol a catena lunga e acidi grassi, includendo così esteri delle vitamine A e D;
- le ceramidi, ossia amidi di acidi grassi con di- o triidrossibasi a catena lunga contenenti 12−22 atomi di carbonio nella catena idrocarburica, di cui un esempio è la sfingosina.
Lipidi complessi
A differenza dei lipidi semplici, i lipidi complessi sono formati da più di due tipi di unità strutturali, e includono tra gli altri:
- i fosfolipidi, ossia esteri del glicerolo con acidi grassi, acido fosforico, e altri gruppi contenenti azoto;
- l’acido fosfatidico, ossia diacilglicerolo esterificato ad acido fosforico;
- la fosfatidilcolina o lecitina, la fosfatidiletanolammina, la fosfatidilserina, e il fosfatidilinositolo, ossia acido fosfatidico legato rispettivamente a colina, etanolammina, serina e inositolo;
- sfingolipidi, derivati delle ceramidi;
- sfingomielina, ossia ceramide fosforilcolina;
- cerebrosidi: sono ceramide monoesosido ossia ceramide legata a una sola unità glucidica al gruppo idrossilico terminale della base;
- ceramide di- e poliesosido, ossia legata rispettivamente a un disaccaride o a un tri- o un oligosaccaride;
- cerebroside solfato, ossia ceramide monoesosido esterificato a un gruppo solfato.
Lipidi derivati
I lipidi derivati sono i mattoni costituenti dei lipidi semplici e complessi e si possono presentare come tali o essere rilasciati dagli altri due gruppi principali a seguito di idrolisi.
Includono gli acidi grassi e alcoli, le vitamine A, D, E e K, gli idrocarburi e gli steroli.
Funzioni dei lipidi
I lipidi svolgono una vasta gamma di funzioni che vanno ben oltre il loro ruolo di riserva energetica. Grazie alla loro eterogeneità chimica e alla coesistenza, in molte molecole lipidiche, di regioni polari e non polari, i lipidi sono particolarmente adatti a partecipare a un’ampia varietà di processi biologici.
A livello cellulare, contribuiscono all’organizzazione, alla stabilità e alle proprietà dinamiche delle membrane, influenzandone la permeabilità, la fluidità e le interazioni con le proteine. Oltre a questi ruoli strutturali, le molecole lipidiche e i loro derivati metabolici sono fondamentali per la segnalazione intracellulare, la comunicazione intercellulare e la regolazione ormonale.
A livello dell’intero organismo, i lipidi sono coinvolti nell’immagazzinamento e utilizzo dell’energia, nella protezione e nell’isolamento dei tessuti, nella digestione e nell’assorbimento dei nutrienti e nell’eliminazione del colesterolo in eccesso. Inoltre, alcuni lipidi partecipano alle interazioni con l’ambiente esterno, influenzando la percezione sensoriale e, in molte specie, mediando la comunicazione chimica.
I principali ruoli funzionali dei lipidi sono descritti di seguito.
| Categoria funzionale | Esempi tipici |
|---|---|
| Accumulo e rilascio di energia | Trigliceridi immagazzinati nel tessuto adiposo; acidi grassi utilizzati come combustibile metabolico |
| Componenti strutturali delle membrane | Fosfolipidi, colesterolo, glicolipidi |
| Molecole regolatorie e di segnalazione | Diacilglicerolo, ceramidi, sfingosina, fattore di attivazione piastrinica |
| Funzione ormonale | Ormoni steroidei (estrogeni, androgeni, cortisolo); eicosanoidi (prostaglandine, leucotrieni, trombossani) |
| Ruoli protettivi e isolanti | Grasso sottocutaneo; cuscinetti adiposi strutturali; guaina mielinica |
| Funzioni digestive e di assorbimento | Sali biliari; lipidi alimentari che facilitano lo svuotamento gastrico e la secrezione pancreatica |
| Funzione escretoria | Eliminazione del colesterolo attraverso i sali biliari e l’escrezione fecale |
| Ruoli ecologici e sensoriali | Feromoni; lipidi che contribuiscono alla consistenza e al sapore degli alimenti |
Energia e nutrizione
Gli acidi grassi, immagazzinati nelle cellule sotto forma di trigliceridi, sono una delle principali fonti di energia nell’uomo. Sono anche il combustibile più ricco di energia, fornendo in media 9 kcal/g, mentre i carboidrati e le proteine forniscono circa 4 kcal/g.
Alcuni lipidi sono nutrienti essenziali. Tra questi vi sono la vitamina A, essenziale per la vista; la vitamina D, essenziale per il metabolismo del calcio; la vitamina E, che previene l’autossidazione dei lipidi insaturi; la vitamina K, essenziale per la normale coagulazione del sangue; e gli acidi grassi essenziali, ovvero l’acido α-linolenico e l’acido linoleico che sono rispettivamente i precursori degli acidi grassi polinsaturi omega-3 e degli omega-6.
Funzioni strutturali e di membrana
I fosfolipidi, il colesterolo e i glicolipidi, insieme alle proteine, sono gli elementi costitutivi fondamentali delle membrane biologiche. Inoltre, i lipidi possono agire come recettori, antigeni e ancore di membrana per le proteine nella membrana plasmatica. Influenzando la struttura della membrana, modulano anche l’attività e la funzionalità degli enzimi ad essa associati.
Funzioni regolatorie e ormonali
Molti lipidi agiscono come regolatori dei processi intracellulari. Ne sono un esempio il diacilglicerolo, le ceramidi, la sfingosina e il fattore di attivazione piastrinica, che partecipano tutti alle vie di segnalazione intracellulari.
Molti ormoni sono lipidi. Gli ormoni steroidei, come gli estrogeni, gli androgeni e il cortisolo, sono sintetizzati a partire dal colesterolo. Al contrario, le prostaglandine, le prostacicline, i leucotrieni, i trombossani e altri composti correlati, noti collettivamente come eicosanoidi, derivano dagli omega-3 e dagli acidi grassi polinsaturi omega-6, come l’acido arachidonico, l’acido eicosapentaenoico e l’acido docosaesaenoico.
Funzioni protettive e isolanti
Alcuni depositi di grasso non vengono mobilizzati durante il digiuno e sono classificati come grasso strutturale. La loro funzione è quella di mantenere gli organi e i nervi nella posizione corretta e di proteggerli da lesioni traumatiche e shock. Esempi tipici sono i cuscinetti adiposi sui palmi delle mani e sulle piante dei piedi, che proteggono le ossa sottostanti dalla pressione meccanica.
Negli esseri umani è presente uno strato sottocutaneo di grasso che isola il corpo, riducendo la perdita di calore e contribuendo al mantenimento della temperatura corporea.
Nell’epidermide, i lipidi svolgono un ruolo chiave nel mantenimento della barriera idrica della pelle.
I lipidi fungono anche da isolanti elettrici degli assoni. Gli assoni sono avvolti ripetutamente dalle membrane plasmatiche delle cellule di Schwann nel sistema nervoso periferico e dagli oligodendrociti nel sistema nervoso centrale. Questo rivestimento multistrato è noto come guaina mielinica.
Funzioni digestive ed escretorie
Nel tratto digestivo, i lipidi facilitano la digestione riducendo la secrezione gastrica, rallentando lo svuotamento gastrico e stimolando la secrezione biliare e pancreatica.
I sali biliari sono detergenti naturali sintetizzati nel fegato e secreti nella bile. Essi solubilizzano i fosfolipidi e il colesterolo nella bile e consentono la secrezione del colesterolo nell’intestino. L’escrezione del colesterolo e dei sali biliari rappresenta la principale via attraverso la quale il colesterolo viene eliminato dall’organismo. I sali biliari svolgono anche un ruolo cruciale nella digestione e nell’assorbimento dei lipidi, nonché nell’assorbimento delle vitamine liposolubili nell’intestino.
Funzioni ecologiche e sensoriali
In molti animali, alcuni lipidi vengono secreti nell’ambiente esterno, dove agiscono come feromoni che attraggono o respingono altri organismi. Infine, i lipidi influenzano la consistenza e il sapore degli alimenti, influendo così sulla loro appetibilità.
Digestione
A differenza della digestione dei carboidrati e dell’assorbimento dei monosaccaridi, nonché della digestione delle proteine e dell’assorbimento degli amminoacidi, la digestione dei lipidi e l’assorbimento dei prodotti della loro digestione sono processi che devono tenere conto della scarsa o nulla solubilità di molte delle molecole coinvolte nell’ambiente acquoso dell’intestino.
La digestione dei lipidi inizia nella bocca, prosegue nello stomaco e si completa nel duodeno, mentre i prodotti della digestione vengono assorbiti principalmente nei segmenti successivi dell’intestino tenue, ossia il digiuno e l’ileo.
I lipidi idrofobici giunti nello stomaco si aggregano in goccioline la cui formazione è favorita, oltre che dall’azione di rimescolamento dello stomaco, anche dagli acidi grassi a catena corta, dagli acidi grassi a catena media e dai diacilgliceroli, molecole anfipatiche liberate per azione delle lipasi linguale e gastrica. Queste molecole, disponendosi con la loro regione idrofila sulla superficie della gocciolina e la coda idrofobica rivolta verso l’interno formato da lipidi non polari, agiscono da emulsionanti creando una regione superficiale idrofilica in grado di interagire stabilmente con l’ambiente acquoso circostante.
Nel duodeno, le goccioline lipidiche sono ulteriormente emulsionate dai sali biliari e dai fosfolipidi presenti nella bile rilasciata dalla colecisti. Questo permette la formazione di goccioline sempre più piccole, il che aumenta la superficie disponibile per l’attacco degli enzimi pancreatici responsabili della digestione dei lipidi.
Assorbimento intestinale
Con l’eccezione degli acidi grassi a catena corta e degli acidi grassi a catena media, l’assorbimento intestinale dei prodotti della digestione lipidica richiede la formazione di strutture in grado di trasportare alla superficie luminale degli enterociti molecole non polari. Le strutture in questione sono le micelle miste, più piccole delle goccioline lipidiche e formate dai sali biliari e dai prodotti della digestione lipidica. Hanno una struttura che ricorda un disco ritagliato da una membrana a doppio strato, dove i sali biliari sono disposti sui bordi di taglio, con la regione idrofilica a contatto con l’ambiente acquoso esterno e la regione idrofobica rivolta verso l’interno non polare.
Grazie alle micelle miste la concentrazione dei lipidi a ridosso della superficie luminale degli enterociti aumenta fino a 1.000 volte, il che ne facilita l’assorbimento. Il gradiente di concentrazione tra la superficie della cellula e il suo interno è mantenuto anche dalla rapida riesterificazione intracellulare in esteri del colesterolo, trigliceridi e fosfolipidi dei lipidi assorbiti.
L’assorbimento dei prodotti della digestione dei lipidi avviene attraverso la diffusione passiva e attraverso il trasporto facilitato mediato da specifici trasportatori proteici.
Bibliografia
- Akoh C.C., Min D.B. Food lipids: chemistry, nutrition, and biotechnology. 4th Edition. Boca Raton: CRC Press, 2017. doi:10.1201/9781315151854
- Chow Ching K. Fatty acids in foods and their health implications. 3rd Edition. CRC Press, 2008.
- Dakal T.C., Xiao F., Bhusal C.K., Sabapathy P.C., Segal R., Chen J., Bai X. Lipids dysregulation in diseases: core concepts, targets and treatment strategies. Lipids Health Dis 2025;24(1):61. doi:10.1186/s12944-024-02425-1
- Guyton A.C., Hall J.E. Textbook of medical physiology. 14th Edition. Philadelphia: Elsevier, 2021.
- Heilman D., Woski S., Voet D., Voet J.G., Pratt C.W. Fundamentals of biochemistry: life at the molecular level. 6th Edition. Wiley, 2023.
- Rosenthal M.D., Glew R.H. Medical biochemistry – Human metabolism in health and disease. John Wiley & Sons, 2009.
- Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 4th Edition. St. Louis: Elsevier, 2018.
Domande Frequenti
Qual è la differenza principale tra lipidi semplici e complessi?
I lipidi semplici sono formati da due soli tipi di unità strutturali, come gli esteri del glicerolo nei trigliceridi. I lipidi complessi, invece, presentano più di due tipi di unità strutturali e includono elementi come acido fosforico o gruppi glucidici, come nei fosfolipidi.
Perché la digestione dei lipidi è più complessa di quella dei carboidrati?
A differenza dei carboidrati solubili, molti lipidi sono idrofobici e insolubili nell'ambiente acquoso intestinale. Per questo motivo la loro digestione richiede un processo di emulsione meccanica ed enzimatica che necessita tassativamente dell'intervento dei sali biliari e delle lipasi.
Quali sono le funzioni biologiche dei lipidi nell'organismo?
Oltre a essere una riserva energetica da 9 kcal/g, i lipidi costituiscono la struttura delle membrane cellulari, agiscono come isolanti termici nel tessuto sottocutaneo ed elettrici nella guaina mielinica. Sono inoltre fondamentali per la segnalazione ormonale e l'assorbimento vitaminico.