Il trealosio è un disaccaride formato da due unità di α-D-glucosio unite da un legame glicosidico α,α-(1→1).[1]
È uno zucchero non riducente, ed è quindi dotato di una bassa reattività chimica. In Natura l’unico altro zucchero non riducente ampiamente distribuito è il saccarosio.[2]
È presente in tutti i principali gruppi di organismi quali batteri, funghi, lieviti, alghe, piante e invertebrati, ma è assente nei vertebrati, che mancano delle specifiche vie biosintetiche.[3]
Le fonti alimentari sono alcune specie di funghi, i lieviti, i crostacei, il miele e i cibi lavorati nei quali sia aggiunto come additivo.[4]
Può svolgere funzioni differenti come deposito di carboidrati, fonte di energia, componente dei lipidi di membrana, molecola di segnalazione tra piante e microorganismi simbionti o patogeni, protezione nei confronti di stress abiotici.[5] In forma fosforilata, nelle piante, è coinvolto nella regolazione del metabolismo dei carboidrati.[6]
I mammiferi utilizzano il trealosio esogeno come fonte di carbonio ed energia. La sua digestione richiede l’intervento dell’enzima trealasi (EC 3.2.1.28), in una reazione che porta alla liberazione delle due molecole di glucosio che lo compongono.[7]
In Sintesi: Punti Chiave
- Proprietà chimiche: un disaccaride non riducente composto da due unità di α-D-glucosio unite da un legame glicosidico α,α-(1→1), caratterizzato da bassa reattività chimica e alta resistenza all’idrolisi.
- Fonti alimentari: presente in piccole quantità in funghi, lieviti, crostacei e miele, oltre che in cibi lavorati in cui viene aggiunto come additivo.
- Digestione e metabolismo: scisso nell’intestino tenue in due molecole di glucosio dalla trealasi, fungendo da fonte di carbonio ed energia.
- Funzioni biologiche: agisce come riserva di carboidrati, fonte di energia, componente lipidico strutturale e protezione contro lo stress abiotico in batteri, funghi, piante e invertebrati.
- Significato clinico: mutazioni nei geni della trealasi causano intolleranza al trealosio, portando a diarrea osmotico-fermentativa e sintomi gastrointestinali.
Indice
- Cenni storici
- Proprietà chimiche
- Fonti alimentari
- Biosintesi
- Trealosio nelle piante
- Funzioni
- Meccanismi molecolari del potenziale protettivo
- Digestione del trealosio
- Deficit della trealasi
- Bibliografia
Cenni storici
È stato scoperto in un fungo della segale cornuta, genere Claviceps, da H.A.L. Wiggers nel 1832.[8] Ventisette anni dopo, nel 1859, M. Berthelot lo isolò da Trehala manna, da cui il nome, una sostanza a forma di bozzolo prodotta dall’attività larvale dei tonchi della famiglia dei Curculionidae su alcune specie di Echinops.[9]
Proprietà chimiche
Il trealosio, al pari dei disaccaridi lattosio, maltosio e saccarosio, ha un peso molecolare di 342,30 g/mol e formula molecolare C12H22O11.
Secondo la nomenclatura IUPAC il suo nome è α-D-gluco-esopiranosile α-D-gluco-esopiranoside.[1]
Poiché il legame glicosidico α,α-(1→1) interessa la funzione aldeidica di due unità di glucosio, ossia i due carboni anomerici, in soluzione non può esistere in una forma a catena aperta con un gruppo aldeidico libero, forma che sarebbe in grado di agire come agente riducente. Ne consegue che il trealosio è uno zucchero non riducente, ed è quindi dotato di una bassa reattività chimica.[10]
A temperatura ambiente si presenta come cristalli ortorombici bianchi.
Il suo punto di fusione è pari a 203 °C (397,4 °F; 476,15 K).
Il trealosio fornisce circa 4 kcal/g.[1]
La sua dolcezza è pari a circa il 45% di quella del saccarosio.[11]
| Categoria | Parametro | Specifiche | Dettagli | Riferimento / Valore |
|---|---|---|---|---|
| Identificativi | Numero CAS | Registro | Riferimento banca dati CAS | 99-20-7 |
| PubChem CID | ID Composto | National Center for Biotechnology Information | 7427 | |
| Caratteristiche chimiche | Formula molecolare | Composizione | Formula elementare | C12H22O11 |
| Peso molecolare | Massa | Massa molare (anidro) | 342.30 g/mol | |
| Caratteristiche strutturali | Legame glicosidico | Tipo di legame | Configurazione anomerica | Legame α,α-(1→1)-glucosidico |
| Classe chimica | Categoria | Classificazione degli zuccheri | Disaccaride non riducente |
Fonti alimentari
Le fonti naturali per l’uomo sono alcune specie di funghi (Amanita spp.), specialmente se giovani, i lieviti, tra gli animali i crostacei, e il miele.[10]
Nel 2000, la FDA lo ha riconosciuto come ingrediente e additivo alimentare consentito e classificato, ad esempio, tra gli esaltatori di sapidità e i conservanti. Poiché è presente in piccole quantità in pochi alimenti non lavorati, i cibi lavorati, come gamberetti surgelati, pesce sottovuoto, cereali per la colazione e prodotti da forno, ne sono diventati la principale fonte.[4]
Biosintesi
Sono state identificate cinque vie di biosintesi nei procarioti, di cui solamente una, che è probabilmente anche la più comune, è presente anche negli eucarioti, si compone di due passaggi ed è descritta di seguito.[12][13]
Nel primo passaggio, catalizzato dalla trealosio-6-fosfato sintasi (EC 2.4.1.15), la condensazione dell’UDP-glucosio e del glucosio-6-fosfato porta alla formazione del trealosio-6-fosfato.
Il trealosio-6-fosfato viene quindi defosforilato a trealosio nella reazione di idrolisi catalizzata dalla trealosio-6-fosfato fosfatasi (EC 3.1.3.12).
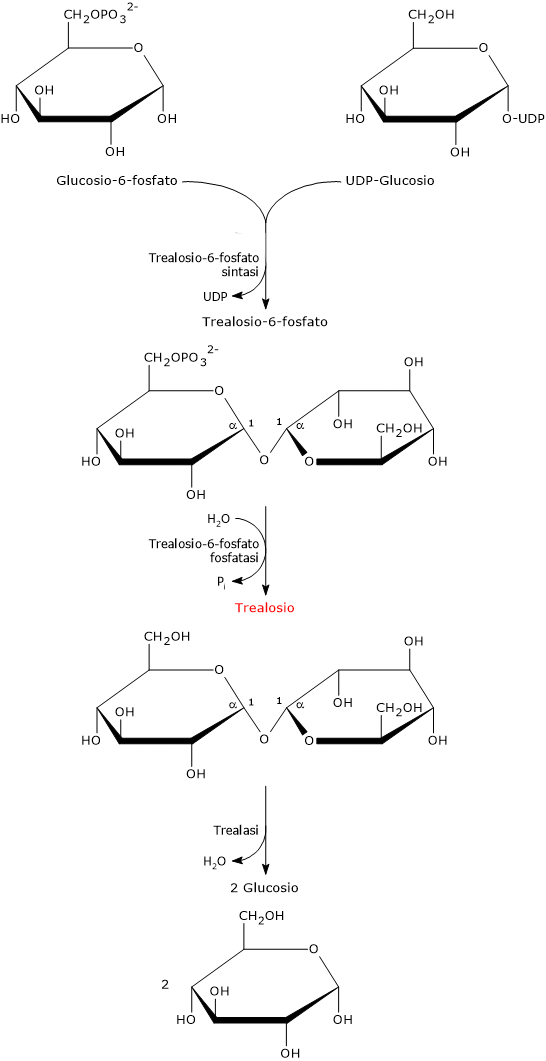
La sua biosintesi è fortemente indotta in seguito all’esposizione a stress abiotici. L’importanza del trealosio è testimoniata anche dal fatto che alcuni organismi possiedono più di una via metabolica per la sua sintesi.[5]
Nessun gene coinvolto nella sua biosintesi o nel suo deposito è presente nei vertebrati, i quali non sembrano aver perso la capacità di produrlo nel corso del processo evolutivo, bensì sembrano non averla mai acquisita.[3]
Trealosio nelle piante
È il principale disaccaride presente nelle piante in crescita.[14]
Nelle piante fu rilevato per la prima volta nel 1913, in Selaginella lepidophylla, nota come pianta della Resurrezione o falsa rosa di Gerico.[15] In seguito fu trovato nelle alghe verdi, nei muschi, nelle felci, e nelle epatiche, piante non-vascolari appartenenti alla divisione Marchantiophyta.[16]
Per molti anni la sua presenza nelle angiosperme fu ritenuta conseguenza di contaminazione microbica o batterica, o un artefatto analitico. Le cose cambiarono quando, dagli anni ’90 dello scorso secolo, in angiosperme resistenti all’essiccamento, come Myrothamnus flabellifolius e Sporobolus spp, ne furono trovate quantità non giustificabili da contaminazioni microbiche.[17][18] Nonostante ciò, nelle angiosperme il trealosio e il suo metabolismo continuarono a essere considerati di scarso interesse fino a che due osservazioni parallele portarono a un cambio di paradigma.
Nel 1998, in Arabidopsis thaliana, una pianta non tollerante all’essiccamento utilizzata come organismo modello in Botanica, sono stati identificati geni codificanti per forme cataliticamente attive della trealosio-6-fosfato sintasi e della trealosio-6-fosfato fosfatasi.[19][20] Inoltre, sempre nelle angiosperme, l’importanza del metabolismo del trealosio è emersa anche a seguito del tentativo di ingegnerizzarne la produzione con l’introduzione di enzimi di origine fungina o batterica. Come risultato sono state ottenute piante transgeniche con un ampio spettro di anomalie fenotipiche, come ritardo nella senescenza e foglie di forma alterata.[21] Queste osservazioni hanno quindi suggerito che nelle angiosperme:
- la capacità di sintetizzare il trealosio non è limitata alle “piante della resurrezione”;
- una perturbazione del suo metabolismo comporta effetti di larga portata sia sul metabolismo stesso che sullo sviluppo.[22]
Attualmente i geni che codificano per la trealosio-6-fosfato sintasi e la trealosio-6-fosfato fosfatasi sono stati identificati in specie di tutti i principali taxa vegetali, il che suggerisce che il metabolismo del disaccaride è probabilmente universale nel regno vegetale.[14]
Funzioni
Il trealosio svolge molteplici funzioni in organismi molto differenti tra loro. Alcune di queste funzioni sono condivise da organismi differenti, mentre altre sono peculiari di specifici organismi.
- Nelle piante, negli insetti, nei nematodi e nei batteri, è coinvolto nella risposta adattativa a stress abiotici, come temperature estreme, variazioni della pressione osmotica e della salinità, privazione di nutrienti o essicazione.[23] Accumulandosi nella cellula, aiuta l’organismo a superare la condizione stressante limitando i danni alle molecole biologiche.[24] Nelle piante il suo accumulo in condizioni di stress è conseguenza dell’attivazione trascrizionale dei geni che codificano per gli enzimi della sua sintesi o dell’inibizione della trealasi.[25] Una volta che la situazione stressante si è risolta il trealosio torna a livelli normali.[6]
- Può fungere da molecola di deposito di carboidrati e trasporto del carbonio. In alcune spore batteriche viene accumulato fino a costituire quasi il 25% del peso secco della spora.[26]
Alcuni batteri possono utilizzarlo come fonte esogena di carbonio.[3]
Nei funghi e lieviti viene conservato durante la fase di dormienza.[27] - Negli insetti è il principale zucchero presente nell’emolinfa, dove rappresenta 80–90% dei carboidrati, e costituisce una fonte di energia rapidamente disponibile per il volo.[16][28]
- È un potenziale metabolita di segnalazione nelle interazioni tra le piante e i microorganismi patogeni e simbionti, nonché con gli insetti erbivori.[14]
- Alcuni batteri e funghi dipendono dal suo metabolismo per la loro infettività.[29][30]
Altre funzioni
È un componente di lipidi presenti nella parete cellulare di specie batteriche appartenenti ai generi Corynebacterium e Mycobacterium, dove svolge un ruolo strutturale.[31] Ad esempio nella parete cellulare di Mycobacterium tuberculosis sono presenti glicolipidi contenenti trealosio, come sulfolipidi, e diacil-, triacil- e poliaciltrealosi, dove gli acidi grassi sono legati ai gruppi idrossilici non del glicerolo ma del disaccaride.[32]
Sembra in grado di proteggere gli acidi grassi insaturi con doppi legami cis dal danno ossidativo.[33] È stato anche proposto che sia in grado di prevenire l’aggregazione delle proteine interagendo con i doppi legami cis delle catene laterali degli aminoacidi aromatici e limitando l’acetilazione dell’ε-amino gruppo della catena laterale di residui di lisina, acetilazione che aumenterebbe l’idrofobicità delle proteine.[33][34]
Ha un ruolo cruciale nella crescita e sviluppo degli insetti, rappresentando in certe fasi del loro sviluppo circa il 20% del totale del pool dei carboidrati.[3]
Il trealosio-6-fosfato è essenziale per lo sviluppo embrionale e vegetativo delle piante, nonché per il metabolismo del saccarosio e dell’amido.[35] E’ stato ipotizzato che la principale funzione del trealosio-6-fosfato sia quella di segnalare e regolare, agendo come un regolatore a feedback negativo, i livelli del saccarosio.[36]
Meccanismi molecolari del potenziale protettivo
Il trealosio interviene nella risposta adattativa a stress abiotici in organismi differenti.
Si ritiene che il suo potenziale protettivo sia dovuto a diversi meccanismi che agiscono in simultanea e sono conseguenza delle sue proprietà chimiche, in particolare quella di essere uno zucchero non riducente.[5]
La sua scarsa reattività chimica assicura che non interagisca facilmente con altre molecole presenti nella cellula, come le proteine, il DNA o l’RNA.[10]
La sua elevata idrofilicità gli permette di formare legami idrogeno con l’acqua più forti rispetto a quelli acqua-acqua. E, sebbene anche altri disaccaridi siano in grado di sostituirsi all’acqua, il suo numero di idratazione è maggiore rispetto a quello degli altri disaccaridi.[11][33] La combinazione della scarsa reattività chimica e dell’elevata idrofilicità lo rendono compatibile con il metabolismo cellulare anche ad alte concentrazioni.[18][37]
La flessibilità del legame glicosidico α,α-(1→1), maggiore rispetto alla flessibilità del legame glicosidico di altri disaccaridi, gli permette di conformarsi più facilmente con i gruppi polari presenti sulle altre molecole.[10]
È importante anche la grande resistenza del legame glicosidico α,α-(1→1) all’idrolisi acida e alla scissione a opera delle glucosidasi.[6]
Tuttavia la differenza chiave che distingue il trealosio dagli altri disaccaridi è la sua capacità di formare una sorta di struttura vetrosa attorno alle molecole che risulterebbe stabile anche alle alte temperature e in caso di essiccamento. Ciò che sembra accadere è che il trealosio, sostituendosi alle molecole d’acqua normalmente legate da legami idrogeno con le molecole biologiche, come ad esempio le proteine le manterrebbe opportunamente ripiegate nella loro struttura nativa.[11]
Digestione del trealosio
Nei mammiferi, la digestione dei carboidrati avviene per la maggior parte nel duodeno. L’alfa-amilasi pancreatica e le idrolasi presenti nell’orletto a spazzola degli enterociti idrolizzano disaccaridi, oligosaccaridi e polisaccaridi nei monosaccaridi costituenti, ovvero glucosio, fruttosio e galattosio. Segue l’assorbimento dei monosaccaridi prodotti.
Il legame glicosidico α,α-(1→1) del trealosio è idrolizzato nella reazione catalizzata dall’enzima trealasi, reazione che porta al rilascio di due molecole di glucosio.[7]
Deficit della trealasi
Nel genoma umano sono presenti due geni che codificano per isoforme della trealasi.[38]
Mutazioni a carico di questi geni portano alla sintesi di enzimi con funzionalità scarsa o assente. In conseguenza di ciò il trealosio non digerito raggiunge la parte distale dell’intestino tenue, dove può causare diarrea osmotica, e, una volta raggiunto il colon, può essere fermentato dai batteri del microbiota intestinale, che è parte del più ampio microbiota umano. Ne risulta la produzione di un eccesso di gas e acidi grassi a catena corta, come l’acido acetico, l’acido propionico e l’acido butirrico, che causano gli altri segni e sintomi della deficienza della trealasi quali diarrea osmotico-fermentativa, malassorbimento e altri sintomi addominali.[39][40]
Poiché non esistono cure per la deficienza della trealasi, l’unico trattamento è evitare o ridurre il consumo di cibi che contengono trealosio.
Bibliografia
- ^ a b c National Center for Biotechnology Information. PubChem Compound Summary for CID 7427, Trehalose. https://pubchem.ncbi.nlm.nih.gov/compound/Trehalose. Accessed Sept. 25, 2025.
- ^ Yoon Y.S., Cho E.D., Jung Ahn W., Won Lee K., Lee S.J., Lee H.J. Is trehalose an autophagic inducer? Unraveling the roles of non-reducing disaccharides on autophagic flux and alpha-synuclein aggregation. Cell Death Dis 2017;8(10):e3091. doi:10.1038/cddis.2017.501
- ^ a b c d Argüelles J.C. Why can’t vertebrates synthesize trehalose? J Mol Evol 2014;79:111-116. doi:10.1007/s00239-014-9645-9
- ^ a b Evaluations of the joint FAO/WHO expert committee on food additives (JECFA). alpha,alpha-Trehalose. Evaluation year: 2000. https://apps.who.int/food-additives-contaminants-jecfa-database/Home/Chemical/4067
- ^ a b c Vanaporn M., Titball R.W. Trehalose and bacterial virulence. Virulence 2020 Dec;11(1):1192-1202. doi:10.1080/21505594.2020.1809326
- ^ a b c Lee H.J., Yoon Y.S., Lee S.J. Mechanism of neuroprotection by trehalose: controversy surrounding autophagy induction. Cell Death Dis 2018;9(7):712. doi:10.1038/s41419-018-0749-9
- ^ a b Rosenthal M.D., Glew R.H. Medical biochemistry: human metabolism in health and disease. A John Wiley & sons, Inc., Publication, 2009.
- ^ Wiggers H.A.L. Untersuchung über das Mutterkorn, Secale cornutum. Annalen der Pharmacie 1832;1(2):129-182. doi:10.1002/jlac.18320010202
- ^ Berthelot M. Nouvelles recherches sur les corps analogues au sucre de canne. Annales de Chimie et de Physique 1859 3a serie;55:269-282.
- ^ a b c Colaco C., Kampinga J., Roser B. Amorphous stability and trehalose. Science 1995;268(5212):788. doi:10.1126/science.7754360
- ^ a b c Jain N.K., Roy I. Effect of trehalose on protein structure. Protein Sci 2009;18(1):24-36. doi:10.1002/pro.3
- ^ Avonce N., Mendoza-Vargas A., Morett E., Iturriaga G. Insights on the evolution of trehalose biosynthesis. BMC Evol Biol 2006;6:109. doi:10.1186/1471-2148-6-109
- ^ Cabib E., Leloir L.F. The biosynthesis of trehalose phosphate. J Biol Chem 1958;231(1):259-75.
- ^ a b c Lunn J.E., Delorge I., Figueroa C.M., Van Dijck P., Stitt M. Trehalose metabolism in plants. Plant J 2014;79(4):544-67. doi:10.1111/tpj.12509
- ^ Anselmino O. and Gilg E. Uber das Vorkommen von Trehalose in Selaginella lepidophylla. Ber Deut Pharm Ges 1913;23:326-330.
- ^ a b Elbein A.D. The metabolism of alpha,alpha-trehalose. Adv Carbohydr Chem Biochem 1974;30:227-56. doi:10.1016/s0065-2318(08)60266-8
- ^ Drennan P.M., Smith M.T., Goldsworthy D., Van Staden J. The occurrence of trehalose in the leaves of the desiccation-tolerant angiosperm Myrothamnus flabellifolius Welw. J Plant Physiol 1993;142(4):493-6. doi:10.1016/S0176-1617(11)81257-5
- ^ Iturriaga G., Gaff D.F., Zentella R. New desiccation-tolerant plants, including a grass, in the central highlands of Mexico, accumulate trehalose. Australian Journal of Botany 2000;48(2):153-8. doi:10.1071/BT98062
- ^ Blazquez M., Santos E., Flores C., Martinez-Zapater J., Salinas J., Gancedo C. Isolation and molecular characterization of the Arabidopsis TPS1 gene, encoding trehalose-6-phosphate synthase. Plant J 1998;13(5):685-9. doi:10.1046/j.1365-313x.1998.00063.x
- ^ Vogel G., Aeschbacher R.A., Muller J., Boller T., Wiemken A. Trehalose-6-phosphate phosphatases from Arabidopsis thaliana: identification by functional complementation of the yeast tps2 mutant. Plant J 1998;13:673-683. doi:10.1046/j.1365-313X.1998.00064.x
- ^ Goddijn O.J., Verwoerd T.C., Voogd E., Krutwagen R.W., de Graaf P.T., van Dun K., Poels J., Ponstein A.S., Damm B., Pen J. Inhibition of trehalase activity enhances trehalose accumulation in transgenic plants. Plant Physiol 1997;113(1):181-90. doi:10.1104/pp.113.1.181
- ^ Figueroa C.M., Lunn J.E. A tale of two sugars: trehalose 6-phosphate and sucrose. Plant Physiol 2016;172(1):7-27. doi:10.1104/pp.16.00417
- ^ Crowe J.H., Hoekstra F.A., Crowe L.M. Anhydrobiosis. Annu Rev Physiol 1992;54:579-99. doi:10.1146/annurev.ph.54.030192.003051
- ^ Richards A.B., Krakowka S., Dexter L.B., Schmid H., Wolterbeek A.P., Waalkens-Berendsen D.H., Shigoyuki A., Kurimoto M. Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies. Food Chem Toxicol 2002;40(7):871-98. doi:10.1016/s0278-6915(02)00011-x
- ^ Fernandez O., Béthencourt L., Quero A., Sangwan R.S., Clément C. Trehalose and plant stress responses: friend or foe? Trends Plant Sci 2010;15(7):409-17. doi:10.1016/j.tplants.2010.04.004
- ^ McBride M.J., Ensign J.C. Effects of intracellular trehalose content on Streptomyces griseus spores. J Bacteriol 1987;169(11):4995-5001. doi:10.1128/jb.169.11.4995-5001.1987
- ^ Barton J.K., Den Hollander J.A., Hopfield J.J., Shulman R.G. 13C nuclear magnetic resonance study of trehalose mobilization in yeast spores. J Bacteriol 1982;151(1):177-85. doi:10.1128/jb.151.1.177-185.1982
- ^ Wyatt G.R., Kale G.F. The chemistry of insect hemolymph. II. Trehalose and other carbohydrates. J Gen Physiol 1957;40(6):833-47. doi:10.1085/jgp.40.6.833
- ^ Wilson R.A., Jenkinson J.M., Gibson R.P., Littlechild J.A., Wang Z.Y., Talbot N.J. Tps1 regulates the pentose phosphate pathway, nitrogen metabolism and fungal virulence. EMBO J 2007 Aug;26(15):3673-85. doi:10.1038/sj.emboj.7601795
- ^ Djonović S., Urbach J.M., Drenkard E., Bush J., Feinbaum R., Ausubel J.L., Traficante D., Risech M., Kocks C., Fischbach M.A., Priebe G.P., Ausubel F.M. Trehalose biosynthesis promotes Pseudomonas aeruginosa pathogenicity in plants. PLoS Pathog 2013;9(3):e1003217. doi:10.1371/journal.ppat.1003217
- ^ Iturriaga G., Suárez R., Nova-Franco B. Trehalose metabolism: from osmoprotection to signaling. Int J Mol Sci 2009;10(9):3793-3810. doi:10.3390/ijms10093793
- ^ Kalscheuer R., Koliwer-Brandl H. Genetics of mycobacterial trehalose metabolism. Microbiol Spectr 2014;2(3). doi:10.1128/microbiolspec.MGM2-0002-2013
- ^ a b c Furuki T., Oku K., Sakurai M. Thermodynamic, hydration and structural characteristics of alpha,alpha-trehalose. Front Biosci (Landmark Ed). 2009;14(9):3523-35. doi:10.2741/3468
- ^ Moruno Algara M., Kuczyńska-Wiśnik D., Dębski J., Stojowska-Swędrzyńska K., Sominka H., Bukrejewska M., Laskowska E. Trehalose protects Escherichia coli against carbon stress manifested by protein acetylation and aggregation. Mol Microbiol 2019;112(3):866-880. doi:10.1111/mmi.14322
- ^ Schluepmann H., Pellny T., van Dijken A., Smeekens S. and Paul M. Trehalose 6-phosphate is indispensable for carbohydrate utilization and growth in Arabidopsis thaliana. Proc Natl Acad Sci 2003;100(11):6849-6854. doi:10.1073/pnas.1132018100
- ^ Yadav U.P., Ivakov A., Feil R., Duan G.Y., Walther D., Giavalisco P., Piques M., Carillo P., Hubberten H.M., Stitt M., Lunn J.E. The sucrose-trehalose 6-phosphate (Tre6P) nexus: specificity and mechanisms of sucrose signalling by Tre6P. J Exp Bot 2014;65(4):1051-68. doi:10.1093/jxb/ert457
- ^ Lin Q., Gong J., Zhang Z., Meng Z., Wang J., Wang S., Sun J., Gu X., Jin Y., Wu T., Yan N., Wang Y., Kai L., Jiang J., Qi S. The Arabidopsis thaliana trehalose-6-phosphate phosphatase gene AtTPPI regulates primary root growth and lateral root elongation. Front Plant Sci 2023;13:1088278. doi:10.3389/fpls.2022.1088278
- ^ TREH trehalase [Homo sapiens]. Gene ID: 11181, updated on 19-Aug-2025. https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=11181
- ^ Holtug K., Clausen M.R., Hove H., Christiansen J., Mortensen P.B. The colon in carbohydrate malabsorption: short-chain fatty acids, pH, and osmotic diarrhoea. Scand J Gastroenterol 1992;27(7):545-52. doi:10.3109/00365529209000118
- ^ Arola H., Koivula T., Karvonen A.L., Jokela H., Ahola T., Isokoski M. Low trehalase activity is associated with abdominal symptoms caused by edible mushrooms. Scand J Gastroenterol 1999;34(9):898-903. doi:10.1080/003655299750025372
Domande Frequenti
Perché il trealosio è considerato uno zucchero non riducente?
Il trealosio è uno zucchero non riducente perché il legame glicosidico α,α-(1→1) interessa direttamente i due carboni anomerici delle unità di glucosio. Bloccando la funzione aldeidica, la molecola non può aprirsi in soluzione per agire come agente riducente.
Come viene digerito il trealosio nell'organismo umano?
Il trealosio viene idrolizzato a livello dell'intestino tenue dall'enzima trealasi, che scinde il legame glicosidico rilasciando due molecole di glucosio liberamente assimilabili. Una carenza di questo enzima può causare intolleranza.
Che ruolo svolge il trealosio nell'emolinfa degli insetti?
Negli insetti il trealosio funge da principale zucchero di trasporto nell'emolinfa. Mobilitato rapidamente dai corpi adiposi, garantisce il fabbisogno energetico immediato richiesto dai muscoli durante il volo prolungato e intenso.