Gli acidi grassi sono acidi carbossilici con una catena idrocarburica di lunghezza variabile. Il gruppo carbossilico e la catena idrocarburica determinano le proprietà chimico-fisiche della molecola.
La catena idrocarburica nella maggior parte dei casi è priva di ramificazioni e generalmente con un numero pari di atomi di carbonio. Può contenere solo legami singoli, nel caso degli acidi grassi saturi, o presentare almeno un doppio/triplo legame, nel caso degli acidi grassi insaturi.
Gli acidi grassi appartengono alla classe di composti noti come lipidi, e possono essere classificati sulla base di criteri chimici e fisiologici.
Svolgono importanti funzioni nella cellula. Possono infatti essere ossidati per fornire energia, sono componenti delle membrane cellulari, e precursori di potenti mediatori lipidici bioattivi.
INDICE
Struttura
Gli acidi grassi sono acidi carbossilici di formula generale R–COOH, dove R– è una catena idrocarburica. Sono quindi formati da soli atomi di carbonio, idrogeno e ossigeno.
Un acido carbossilico è un composto organico che presenta il gruppo funzionale –COOH, o gruppo carbossilico, nel quale un atomo di carbonio lega tramite doppio legame un atomo di ossigeno a formare un gruppo carbonilico, e tramite legame semplice un gruppo ossidrilico.
Il gruppo R–, che impegna il quarto legame del carbonio carbossilico, nel caso più semplice è un atomo di idrogeno, e l’acido carbossilico corrispondente è l’acido formico, oppure è una catena idrocarburica che a sua volta potrà essere:
- lineare o ramificata;
- con unità carbocicliche;
- con un numero pari o dispari di atomi di carbonio;
- priva di doppi/tripli legami, dunque satura;
- con doppi/tripli legami, dunque insatura.
Gli acidi grassi presenti negli alimenti hanno generalmente una catena idrocarburica lineare, con un numero pari di atomi di carbonio, una lunghezza compresa tra 4 e 24 atomi, catena che può essere satura o presentare uno o più doppi legami. Per una lista di quelli più importanti per l’alimentazione dell’uomo si veda la pagina: elenco degli acidi grassi.
Gli acidi grassi ramificati sono comuni nei batteri Gram-positivi, sono presenti in basse concentrazioni nei lipidi del latte e della carne dei ruminanti, come ad esempio l’acido fitanico, mentre sono rari nei lipidi di origine vegetale.
Polarità
Il gruppo carbossilico è un gruppo acido polare, mentre la catena idrocarburica ha carattere non polare. L’idrofobicità della catena aumenta all’aumentare della lunghezza, e questo determina il grado di solubilità dell’acido grasso in solventi polari e non polari.
Negli acidi grassi a catena corta, la polarità del gruppo carbossilico vince sulla idrofobicità della catena, la molecola presenta carattere polare e risulta solubile in solventi polari come l’acqua e l’etanolo. L’acido butirrico è un esempio di acido grasso solubile in solventi polari. A partire dall’acido caproico, la cui catena carboniosa è di due atomi più lunga rispetto a quella dell’acido butirrico, la polarità va gradualmente diminuendo passando dall’acido caprilico, all’acido caprico e all’acido laurico, che hanno catene carboniose rispettivamente composte da 8, 10, e 12 atomi.
Con catene superiori ai 16-18 atomi di carbonio, quindi dall’acido palmitico e acido stearico in avanti, le molecole risultano del tutto insolubili solventi polari. Risultano quindi insolubili in solventi polari anche l’acido arachidico, l’acido beenico e l’acido lignocerico, tre molecole sature con catene rispettivamente di 20, 22 e 24 atomi di carbonio.
Punto di fusione ed ebollizione
Al pari della polarità, anche il punto di fusione e il punto di ebollizione sono influenzati dalle proprietà della catena carboniosa.
I punti di fusione e di ebollizione degli acidi grassi saturi aumentano all’aumentare del peso molecolare, e quindi della lunghezza della catena.
Le catene non ramificate prive di doppi legami hanno un andamento relativamente lineare, anche se un po’ a zig-zag. Ciò consente alle molecole di impacchettarsi in modo ravvicinato, il che permette lo stabilirsi di numerosi legami idrofobici intercatena che stabilizzano la struttura in una disposizione quasi cristallina. Questo determina l’innalzamento del punto di fusione e di ebollizione.
I punti di fusione ed ebollizione degli acidi grassi insaturi sono più bassi rispetto ai corrispondenti acidi grassi saturi. Ciò è conseguenza di differenze nella geometria molecolare della catena carboniosa dovute alla presenza di doppi legami.
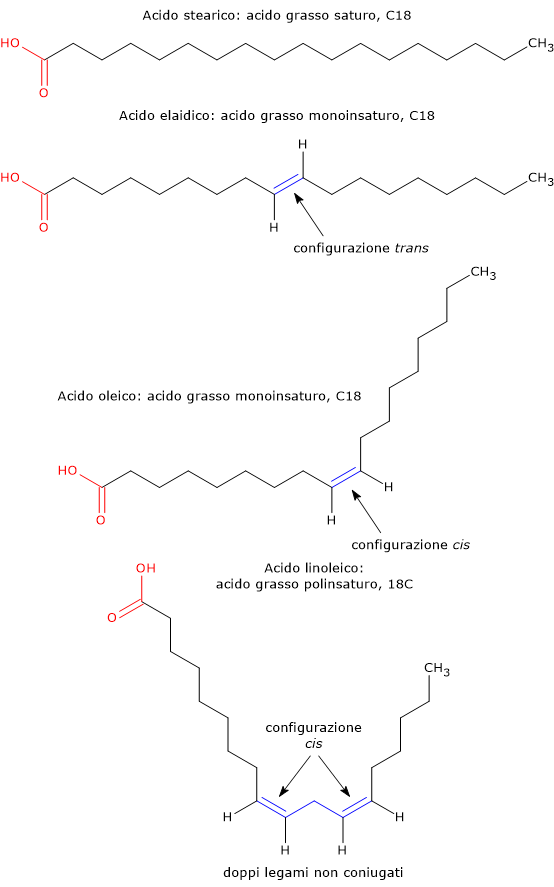 Il doppio legame ha una struttura planare rigida, ossia non permette rotazioni, possibili invece nel legame singolo, e può essere considerato come un piano al quale la catena carboniosa si attacca e procede, o entra ed esce. Quando in una molecola è presente almeno una struttura rigida, che può essere anche un anello, la molecola può presentare isomeria geometrica o isomeria cis–trans.
Il doppio legame ha una struttura planare rigida, ossia non permette rotazioni, possibili invece nel legame singolo, e può essere considerato come un piano al quale la catena carboniosa si attacca e procede, o entra ed esce. Quando in una molecola è presente almeno una struttura rigida, che può essere anche un anello, la molecola può presentare isomeria geometrica o isomeria cis–trans.
Se l’ingresso e l’uscita della catena avvengono sullo stesso lato del piano, il doppio legame è detto in configurazione cis, mentre se l’ingresso e l’uscita avvengono su lati opposto del piano il doppio legame è detto in configurazione trans.
La configurazione cis impone una curvatura nella catena. La catena curva fa si che le molecole si impacchettino con una efficienza minore rispetto agli acidi grassi saturi, stabilendo meno interazioni idrofobiche intermolecolari. Gli aggregati saranno meno stabili e sarà richiesta meno energia per allontanare le molecole le une dalle altre. Ne consegue che sia il punto di fusione che quello di ebollizione degli acidi grassi cis sarà più basso rispetto a quello degli acidi grassi saturi corrispondenti.
La configurazione trans ha una geometria simile a quella del legame singolo, raddrizza la catena carboniosa impartendo all’acido grasso una conformazione simile a quella degli acidi grassi saturi. Le molecole che presentano solo doppi legami trans potranno quindi impacchettarsi con una efficienza simile a quella degli acidi grassi saturi, con i quali avranno punti di fusione ed ebollizione simili.
Le differenze nella geometria tra i doppi legami cis e trans e tra acidi grassi saturi e insaturi influenzano anche le loro funzioni biologiche; ad esempio, gli acidi grassi trans e quelli saturi formano strutture più rigide rispetto ai cis, e questo influenza la fluidità delle membrane biologiche in cui sono presenti.
Classificazione
Esistono diverse modalità di classificazione degli acidi grassi.
Dal punto di vista chimico possono essere classificati sulla base del numero pari o dispari degli atomi di carbonio della catena, sulla presenza o meno di ramificazioni o strutture cicliche, sulla lunghezza della catena, sulla presenza o meno di doppi/tripli legami, sull’isomeria geometrica dei doppi legami, sul numero dei legami doppi/tripli, sulla posizione del primo doppio legame rispetto all’estremità metilica, o se sono presenti in forma libera o esterificata.
Dal unto di vista fisiologico sono classificati sulla base della loro essenzialità per gli esseri umani.
Lunghezza della catena carboniosa
Negli acidi grassi a catena lineare, la catena ha un lunghezza variabile da 1 fino a 30 e più atomi di carbonio.
Sulla base della lunghezza della catena possono essere classificati in acidi grassi:
- a catena corta, nei quali la lunghezza della catena varia da 1 a 5 atomi di carbonio;
- a catena media, dove la catena varia da 6 a 12 atomi di carbonio;
- a catena lunga, dove la catene ha lunghezza compresa da 13 a 21 atomi di carbonio;
- a catena molto lunga, dove la catena carboniosa ha lunghezza pari o superiore a 22 atomi di carbonio.
Saturi e insaturi
In base alla presenza o meno di doppio/tripli legami nella catena carboniosa, possono essere classificati in:
- saturi, che sono privi di doppio/tripli legami;
- insaturi, nei quali è presente almeno un doppio/triplo legame.
In base al numero di doppi/tripli legami nella catena carboniosa, possono essere suddividi in:
- monoinsaturi, quando è presente un solo doppio/triplo legame;
- polinsaturi, quando è presente più di un doppio/triplo legame.
Gli acidi grassi insaturi possono essere classificati in base alla posizione del primo doppio legame rispetto all’estremità metilica della catena.
- Acidi grassi polinsaturi omega-3, quando il primo doppio legame si trova a tre atomi di carbonio di distanza dall’estremità metilica. Esempi sono l’acido alfa-linolenico, l’acido stearidonico, l’acido eicosapentaenoico o EPA e l’acido docosaesaenoico o DHA.
- Acidi grassi polinsaturi omega-6, quando il primo doppio legame si trova a sei atomi di carbonio di distanza dall’estremità metilica, come nell’acido linoleico, nell’acido gamma-linolenico, nell’acido diomo-gamma-linolenico, nell’acido arachidonico e nell’acido adrenico.
- Acidi grassi omega-7, quando il primo doppio legame si trova a sette atomi di carbonio di distanza dall’estremità metilica, come nell’acido palmitoleico.
- Acidi grassi omega-9, quando il primo doppio legame si trova a nove atomi di carbonio di distanza dall’estremità metilica. Esempio sono l’acido oleico, l’acido erucico, l’acido nervonico e l’acido Mead.
- Acidi grassi omega-11, quando il primo doppio legame si trova a undici atomi di carbonio di distanza dall’estremità metilica, come nell’acido gadoleico.
Quando nella catena carboniosa è presente uno o più tripli legami l’acido grasso è definito acetilenico.
I polinsaturi possono essere classificati anche sulla base della presenza di sistemi di doppi legami coniugati.
- Acidi grassi coniugati, quando due o più doppi legami non sono separati da uno o più gruppi metilenici (–CH2–).
- Acidi grassi non coniugati, quando i doppi legami sono separati da almeno un gruppo metilenico. I principali acidi grassi polinsaturi hanno doppi legami non coniugati.
Isomeri cis e trans
Gli acidi grassi insaturi contenenti doppi legami possono essere classificati, sulla base della isomeria geometrica del doppio legame, in:
- acidi grassi cis, i più comuni in Natura e negli alimenti;
- acidi grassi trans o semplicemente grassi trans, meno comuni.
L’acido oleico e l’acido elaidico o elaidinico sono un esempio di isomeria cis–trans: l’acido elaidico è l’isomero trans dell’acido oleico. Altri grassi trans che si ritrovano nei cibi sono l’acido vaccenico e l’acido brassidico.
Acidi grassi ciclici
Contengono una unità ciclica con tre, cinque, come prostaglandine, o anche sei atomi di carbonio.
Acidi grassi esterificati e non esterificati
In natura gli acidi grassi raramente si trovano in forma libera, e in quel caso si parla di acidi grassi non esterificati o liberi, indicati anche come NEFAs o FFAs, acronimi rispettivamente di nonesterified fatty acids e free fatty acids. Nell’uomo ne è presente una piccola quantità nel sangue, derivante da processi di lipolisi, di cui circa tre quarti sono legati all’albumina, mentre il terzo restante è trasportato dalle lipoproteine.
Per la maggior parte sono invece legati tramite legame estere ad altre molecole organiche, quali il glicerolo, il glicerolo-3-fosfato e gli steroli, a formare lipidi più complessi, rispettivamente i trigliceridi, i fosfolipidi, e gli esteri degli steroli, ad esempio gli esteri del colesterolo.
Acidi grassi essenziali
Gli acidi grassi possono essere classificati su base fisiologica, considerando la capacità o meno dell’organismo di sintetizzarli. L’acido alfa-linolenico e l’acido linoleico sono classificati come acidi grassi essenziali, in quanto gli animali non hanno il corredo enzimatico necessario per la loro sintesi, mancando della delta-12 desaturasi (EC 1.14.19.6) e della delta-15 desaturasi (EC 1.14.19.13). Pertanto debbono essere assunti con l’alimentazione.
Funzioni
Nelle cellule svolgono molte funzioni.
- Sono una fonte di energia.
Dopo essere stati rilasciati dai trigliceridi intracellulari, vengono ossidati per produrre ATP all’interno della cellula stessa o, se rilasciati dai trigliceridi del tessuto adiposo, in cellule di altri tessuti e organi. Fegato, cuore e muscolo scheletrico sono i principali siti di ossidazione.
Dalla loro ossidazione le cellule ottengono una quantità di energia maggiore rispetto a quella ottenuta dall’ossidazione di carboidrati e proteine: in media 9 kcal/g contro 4 kcal/g di proteine e carboidrati. Va tuttavia ricordato che dall’ossidazione degli acidi grassi saturi a catena corta si ottiene un quantitativo di energia inferiore: ad esempio dall’acido acetico 3,5 kcal/g, dall’acido propionico 5,0 kcal/g, dall’acido butirrico 6,0 kcal/g, e dall’acido caproico 7,5 kcal/g. - Sono precursori di potenti mediatori lipidici bioattivi che agiscono come parte di vie di trasduzione del segnale. Infatti, gli acidi grassi insaturi come l’acido arachidonico e l’acido docosaesaenoico, derivanti dal flusso sanguigno o rilasciati dai fosfolipidi di membrana, possono essere metabolizzati a potenti mediatori lipidici ad azione antiinfiammatoria, come Lipossine, Maresine, Resolvine della serie D, e a mediatori lipidici proinfiammatori, come Prostaglandine e Leucotrieni.
- Hanno un ruolo strutturale nella formazione delle membrane cellulari essendo un componente dei fosfolipidi che sono a loro volta componenti chiave di tutte le membrane biologiche. E, a differenza delle proteine e degli acidi nucleici, hanno la capacità di essere incorporati intatti nei tessuti, alterando così la composizione acilica dei tessuti stessi.
Bibliografia
- Akoh C.C. and Min D.B. Food lipids: chemistry, nutrition, and biotechnology. 3th Edition. CRC Press, Taylor & Francis Group, 2008
- Burr G. and Burr M. A new deficiency disease produced by the rigid exclusion of fat from the diet. J Biol Chem 1929;82:345-367. doi:10.1016/S0021-9258(20)78281-5
- Calder P.C. Very long-chain n-3 fatty acids and human health: fact, fiction and the future. Proc Nutr Soc 2018;77(1):52-72. doi:10.1017/S0029665117003950
- Chow Ching K. Fatty acids in foods and their health implication. 3th Edition. CRC Press, Taylor & Francis Group, 2008
- Evans H. M. and G. O. Burr. A new dietary deficiency with highly purified diets. III. The beneficial effect of fat in the diet. Proc Soc Exp Biol Med 1928;25:390-397. doi:10.3181/00379727-25-3867
- Mozaffarian D., Katan M.B., Ascherio A., Stampfer M.J., Willett W.C. Trans fatty acids and cardiovascular disease. N Engl J Med 2006;354:1601-1613. doi:10.1056/NEJMra054035
- Petty H.R. Molecular biology of membranes: structure and function. Plenum Press, New York, 1993