Il galattosio è un monosaccaride a sei atomi di carbonio.
Può esistere in forma a catena aperta e in forma ciclica. La forma a catena aperta presenta un gruppo carbonilico a una estremità della catena, il che ne fa un derivato aldeidico, e quindi un aldoesoso e uno zucchero riducente. In forma ciclica può esistere in forma di quattro isomeri differenti.[1]
La principale fonte è il disaccaride lattosio, che è formato da una molecola di galattosio e una di glucosio unite da un legame glicosidico β-(1→4). Nell’intestino tenue il legame glicosidico del disaccaride è scisso nella reazione catalizzata dalla lattasi (EC 3.2.1.108) a dare glucosio e galattosio, che sono quindi assorbiti.[2]
Il galattosio esercita un’ampia gamma di funzioni. Previa conversione in UDP-galattosio e UDP-glucosio attraverso la via di Leloir, può essere utilizzato sia a scopi anabolici che catabolici, a seconda del tipo di tessuto e dello stato energetico della cellula.[3][4]
Un difetto a carico di uno degli enzimi della via di Leloir porta a un accumulo anomalo in molti organi e tessuti di galattitolo e galattonato, molecole derivanti dal galattosio, e causa la galattosemia.[5]
In Sintesi: Punti Chiave
- Proprietà chimiche: monosaccaride a sei atomi di carbonio che può esistere in forma a catena aperta e in quattro forme anomeriche cicliche.
- Fonti alimentari: derivato principalmente dal lattosio presente nel latte e nei latticini; presente in piccole quantità nella frutta, nella verdura e nei legumi.
- Assorbimento e metabolismo: assorbito nell’intestino tenue; metabolizzato principalmente attraverso la via di Leloir; vie alternative includono la via dei polioli e l’ossidazione.
- Funzioni biologiche: essenziale per la sintesi di glicoproteine, glicolipidi e lattosio; contribuisce anche alla determinazione degli antigeni dei gruppi sanguigni ABO.
- Significato clinico: difetti nel suo metabolismo portano alla galattosemia.
Indice
- Cenni storici
- Proprietà chimiche
- Fonti alimentari
- Assorbimento intestinale
- Metabolismo del galattosio
- Galattosemia
- Bibliografia
Cenni storici
Il galattosio fu isolato per la prima volta da Pasteur dal latte nel 1855, e la sua struttura fu chiarita trent’anni più tardi da Kiliani e Fischer. Nel 1860 Berthelot lo chiamò glucosio lattico o galattosio, dal greco galaktos, che significa latte, seguito dal suffisso per gli zuccheri –osio.[6][7]
La comprensione del suo metabolismo ha fatto un significativo passo avanti alla fine degli anni ’40 grazie a Leloir, che identificò gli intermedi zucchero-nucleotide e gli enzimi specifici che convertono il galattosio in glucosio, una scoperta che gli valse il Premio Nobel per la Chimica nel 1970.[8]
Proprietà chimiche
Come tutti gli altri esosi, quali il glucosio e il fruttosio, ha formula molecolare C6H12O6 e massa molare di 180,156 g/mol.
Ha una dolcezza pari al 33% di quella del saccarosio, ed è dolce circa la metà rispetto al glucosio.[9]
| Zucchero (Esoso) | Massa Molare | Dolcezza Relativa (Saccarosio = 100%) |
|---|---|---|
| Saccarosio | 342,296 g/mol | Riferimento standard pari al 100% del potere dolcificante. |
| Glucosio | 180,156 g/mol | Dolcezza intermedia, pari a circa il 75% del saccarosio. |
| Galattosio | 180,156 g/mol | Dolcezza ridotta, pari al 33% del saccarosio (metà del glucosio). |
| Fruttosio | 180,156 g/mol | Dolcezza elevata, pari a circa il 173% rispetto al riferimento. |
Al pari degli altri aldoesosi, ha due enantiomeri che, in base alla convenzione di Fischer o sistema D–L sono detti D-galattosio e L-galattosio. Quest’ultimo non è presente naturalmente negli organismi superiori in quanto non può essere metabolizzato. Pertanto, con il termine di galattosio ci si riferirà all’enantiomero D.
Può esistere in forma ciclica e a catena aperta. La forma a catena aperta, che può essere rappresentata ricorrendo alle proiezioni di Fischer, ha un gruppo aldeidico, e quindi un gruppo carbonilico, a una estremità della molecola.
Il gruppo carbonilico fa sì che il galattosio, al pari degli altri monosaccaridi, e del lattosio e maltosio tra i disaccaridi, sia uno zucchero riducente.[1]
Struttura
La forma a catena aperta è termodinamicamente instabile e in soluzione è presente in tracce.
Poiché le aldeidi possono reagire con l’alcol, la forma a catena aperta del galattosio può ciclizzare a seguito dell’attacco nucleofilo dell’atomo di ossigeno di uno dei gruppi ossidrilici al carbonio carbonilico a formare un emiacetale ciclico.[10]
I composti ciclici con anelli a tre o quattro atomi sono molto meno stabili di quelli a cinque e, soprattutto, a sei atomi, a causa delle elevate interazioni steriche presenti, in analogia a quanto accade con gli alcani ciclici, come il ciclopropano e il ciclobutano, meno stabili del ciclopentano e cicloesano.
Nella forma ciclica, il galattosio presenta quattro stereoisomeri, due con un anello a cinque atomi, quattro atomi di carbonio e uno di ossigeno, ossia un anello furanosico, e due con un anello a sei atomi, cinque di carbonio e uno di ossigeno, quindi un anello piranosico.[11]
In soluzione, le forme piranosiche e furanosiche sono in equilibrio con la forma a catena aperta ma, poiché la forma furanosica è meno stabile della piranosica, l’equilibrio è fortemente spostato verso quest’ultima.[1]
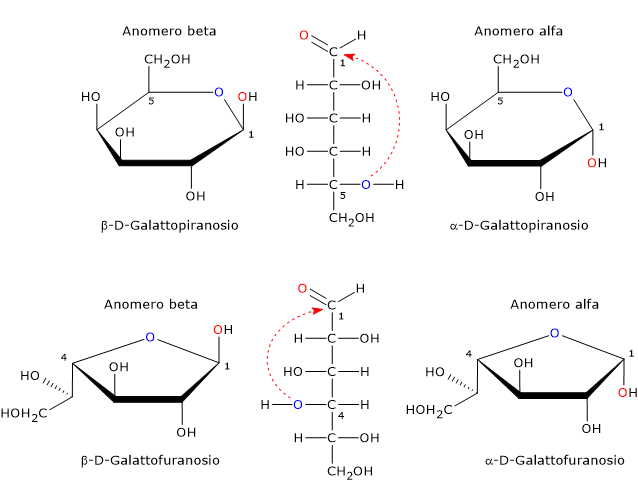
Nei mammiferi è presente la sola forma piranosica, che è un epimero del glucosio differendone solamente per la configurazione del gruppo ossidrilico sul C4. La forma furanosica si trova in molti organismi inferiori, sia procarioti che eucarioti, come batteri, alghe verdi, funghi, protozoi, come Tripanosomatidi, spugne e stelle marine. La forma furanosica si trova anche in specie patogene dove può svolgere un ruolo importante, ad esempio per la sopravvivenza, mentre la sua via biosintetica può essere un bersaglio per lo sviluppo di nuovi farmaci per il trattamento delle infezioni causate dai patogeni stessi.[11]
Anomeri
Il carbonio emiacetalico, un carbonio tetraedrico che lega quattro ligandi differenti, è un centro di chiralità.
Poiché l’attacco nucleofilo dell’atomo di ossigeno può avvenire da entrambi i lati del piano sp2 del carbonio carbonilico, la chiusura dell’anello produce due coppie di anomeri (α e β), sia piranosici che furanosici. In soluzione, queste forme si interconvertono continuamente attraverso la forma a catena aperta fino al raggiungimento dell’equilibrio, un processo noto come mutarotazione.
Tali epimeri differiscono nella configurazione del carbonio emiacetalico e sono detti anomero α e anomero β, mentre il carbonio emiacetalico diventa il carbonio anomerico o centro anomerico.
Si avrà l’anomero α quando l’attacco nucleofilo avviene da sopra rispetto al piano sp2, e in questo caso il gruppo ossidrilico legato al C1 si trova al di sotto del piano dell’anello, e viceversa si avrà l’anomero β, nel quale il gruppo ossidrilico si trova al di sopra del piano dell’anello.
Dunque, se si considerano i possibili anomeri e la forma a catena aperta, il galattosio può esistere in cinque forme differenti (quattro cicliche e una a catena aperta): α-D-galattopiranosio e β-D-galattopiranosio, α-D-galattofuranosio e β-D-galattofuranosio, e la forma a catena aperta.[10]
Nota: l’anomeria è un tipo di isomeria ottica caratteristica dei monosaccaridi.
Fonti alimentari
La maggior parte del galattosio presente nella dieta deriva dal latte e derivati, dove si trova principalmente nella molecola del lattosio, e, in misura minore, come componente di carboidrati più complessi, quali oligosaccaridi e polisaccaridi, detti glicani, componenti a loro volta dei glicolipidi e glicoproteine del latte.[12]
È presente anche in molti altri alimenti, quali frutta, verdura, cereali, legumi, noci e miele, dove si ritrova in forma libera, legato a polifenoli quali flavonoli e antociani, o come componente di glicoproteine e glicolipidi. Nella forma legata è spesso impegnato in legami resistenti all’attacco delle idrolasi intestinali. È il caso dei legami α-galattosidici presenti in oligosaccaridi come il raffinosio e lo stachiosio, tipici dei legumi. Poiché l’intestino umano è privo di α-galattosidasi, enzimi in grado di scindere tali legami, queste molecole raggiungono indigerite il colon, dove sono fermentate dai batteri del microbiota intestinale. Questo processo libera gas che possono concorrere allo sviluppo di discomfort addominale.[13][14]
In forma libera è presente in quantità dell’ordine dei milligrammi. Di seguito alcuni esempi:
| Alimento | Contenuto (mg/100 g parte edibile) | Rilevanza Clinica nella Galattosemia |
|---|---|---|
| Carciofi, funghi, olive, arachidi | Inferiore a 0,1 mg | Apporto infinitesimale e generalmente trascurabile nella dieta. |
| Peperoni, datteri, papaia, anguria, pomodoro | Superiore a 10 mg | Apporto moderato che richiede attenzione nel calcolo giornaliero. |
| Cachi | Fino a 35,4 mg | Apporto elevato che richiede stretto monitoraggio clinico. |
Sebbene siano valori molto bassi devono essere presi in considerazione in caso di galattosemia.[15]
Assorbimento intestinale
Nei mammiferi la digestione dei carboidrati inizia nella bocca e termina nel duodeno e digiuno, dove idrolasi quali la alfa-amilasi pancreatica e le disaccaridasi presenti nella membrana dell’orletto a spazzola degli enterociti idrolizzano disaccaridi, oligosaccaridi e polisaccaridi nei monosaccaridi costituenti, quindi glucosio, fruttosio e galattosio.[2]
L’idrolisi del lattosio a dare glucosio e β-galattosio è catalizzata dalla disaccaridasi lattasi o lattasi-florizin idrolasi, enzima che presenta due siti attivi. Un sito attivo idrolizza il legame glicosidico β-(1→4), dunque una β-glucosidasi, mentre l’altro ha attività florizin-idrolasica, essendo in grado di idrolizzare la florizina e i glicolipidi, come le ceramidi a dare sfingosina e acidi grassi.[16]
Anche la β-galattosidasi batterica (EC 3.2.1.23) dello yogurt è in grado di idrolizzare il lattosio nei monosaccaridi costituenti.[17]
L’assorbimento dei monosaccaridi glucosio e galattosio avviene nell’intestino tenue ed è mediato dal trasportatore SGLT1, una proteina integrale di membrana che opera un trasporto attivo sfruttando l’energia del gradiente elettrochimico transmembrana del sodio.[18]
Il galattosio lascia l’enterocita a livello della membrana basolaterale per diffusione facilitata mediata dal trasportatore GLUT2. Una volta in circolo viene veicolato dal sistema portale al fegato, il principale sito per il suo metabolismo, che ne assorbe quasi il 90%. L’ingresso negli epatociti è mediato da GLUT2.[18]
Il galattosio non assorbito dal fegato potrà raggiungere altri organi, come la ghiandola mammaria in fase di allattamento.
In condizioni fisiologiche si può osservare un aumento della concentrazione ematica del galattosio a seguito dell’assunzione di alcol, che ne riduce l’assorbimento intestinale e il successivo metabolismo epatico.
Metabolismo del galattosio
In assenza di lattosio o altre fonti alimentari di galattosio, il monosaccaride può essere sintetizzato a partire dal glucosio, in quanto la reazione catalizzata dalla UDP-galattosio 4-epimerasi (EC 5.1.3.2), uno dei quattro enzimi della via di Leloir, può procedere in entrambe le direzioni.[2]
In aggiunta alla sintesi de novo e all’apporto con la dieta è molto importante il galattosio derivante dal turnover delle glicoproteine e glicolipidi.[19]
Il galattosio può entrare in tre vie metaboliche principali, la più importante delle quali è la via di Leloir, grazie alla quale può essere convertito in glucosio. Le altre due vie, una ossidativa che porta alla formazione del galattonato e una riduttiva che porta alla conversione del galattosio in galattitolo, in condizioni fisiologiche hanno un ruolo marginale, mentre divengono importanti quando i livelli intracellulari di galattosio sono molto elevati, come nella galattosemia.[4]
Via di Leloir
La via di Leloir, scoperta nel 1948 da Luis Federico Leloir e colleghi, attraverso quattro reazioni enzimatiche porta all’inversione della configurazione del gruppo ossidrilico legato al C-4 del galattosio a dare glucosio.[3]
- Il β-galattosio, un prodotto della digestione del lattosio, viene convertito nell’anomero α nella reazione catalizzata dalla galattosio mutarotasi (EC 5.1.3.3).[20]
- Nel passaggio successivo la galattochinasi (EC 2.7.1.6) catalizza la fosforilazione del galattosio a galattosio-1-fosfato. La fosforilazione blocca all’interno della cellula il monosaccaride e ne mantiene bassa la concentrazione intracellulare, favorendone il trasporto all’interno della cellula.[21]
- Il galattosio-1-fosfato viene attivato a UDP-galattosio nella reazione catalizzata dalla galattosio-1-fosfato uridiltransferasi (EC 2.7.7.12).
- Infine si verifica l’epimerizzazione dell’UDP-galattosio a UDP-glucosio nella reazione catalizzata dalla UDP-galattosio 4-epimerasi.[21]
La via di Leloir permette l’attivazione del galattosio a UDP-galattosio che può essere utilizzato nelle reazioni di glicosilazione di lipidi e proteine.[4]
Questo consente al galattosio di svolgere un ruolo strutturale chiave nelle prime fasi dello sviluppo, e di essere un componente degli antigeni presenti su globuli rossi, in particolare degli antigeni del gruppo ABO.
Nella ghiandola mammaria in allattamento l’UDP-galattosio è un precursore per la sintesi del lattosio, costituendo circa il 30% del galattosio presente nel disaccaride, e viene utilizzato anche nelle reazioni di glicosilazione delle proteine e lipidi del latte stesso.[2]
Destino metabolico dell’UDP-glucosio
L’UDP-glucosio, a seconda del tipo di tessuto e della carica energetica della cellula, può:
- essere utilizzato nelle reazioni di glicosilazione;
- nella sintesi del glicogeno, il principale destino metabolico del galattosio nel fegato;
- entrare nella glicolisi per essere utilizzato a fini energetici;
- entrare nella gluconeogenesi, previa conversione in glucosio-1-fosfato nella reazione catalizzata dalla UDP-glucosio pirofosforilasi (EC 2.7.7.9), e successiva isomerizzazione a glucosio-6-fosfato nella reazione catalizzata dalla fosfoglucomutasi (EC 5.4.2.2).[4][19]
Poiché la UDP-glucosio pirofosforilasi catalizza una reazione reversibile, la sintesi dell’UDP-galattosio dal galattosio-1-fosfato e UTP potrebbe rappresentare una via metabolica alternativa nei soggetti con deficienza di galattosio-1-fosfato uridiltransferasi.[22]
Galattitolo
Una via metabolica alternativa alla via di Leloir porta alla riduzione del galattosio a galattitolo nella reazione catalizzata dalla aldoso reduttasi (EC 1.1.1.21), una reduttasi NADPH-dipendente dotata di un’ampia specificità per i monosaccaridi, essendo in grado di ridurre anche il glucosio a sorbitolo.[19]
L’enzima ha una bassa affinità per il galattosio per cui in condizioni fisiologiche il galattitolo è presente in tracce mentre, nel caso in cui il galattosio si accumuli in quantità elevate, come nella galattosemia, la sua produzione aumenta.
Mentre il sorbitolo può essere convertito a fruttosio nella reazione catalizzata dalla sorbitolo deidrogenasi (EC 1.1.99.21), il galattitolo non è ulteriormente metabolizzato e si accumula nella cellula in quanto scarsamente liposolubile, quindi non in grado di attraversare la membrana plasmatica.[4]
Nota: la via metabolica che porta alla conversione del glucosio in fruttosio è chiamata via dei polioli.
Galattonato
La seconda via metabolica alternativa alla via di Leloir porta all’ossidazione del galattosio a galattonato.[19]
- Nel primo passaggio il galattosio è ossidato a galattono-1,4-lattone, nella reazione catalizzata dalla galattosio deidrogenasi (EC 1.1.1.48); nella reazione il NAD+ funge da agente ossidante e si riduce.
- Nel secondo passaggio il legame lattonico viene idrolizzato, in una reazione che può procedere spontaneamente o essere catalizzata da una lattonasi (EC 3.1.1.25), a dare il galattonato il quale può essere escreto con le urine o ulteriormente ossidato a β-cheto-galattonato nella reazione catalizzata dalla β-L-idrossiacido deidrogenasi, reazione che porta alla produzione di NADH.
- Nell’ultimo passaggio il β-cheto-galattonato viene decarbossilato a xilulosio, in una reazione enzimatica di cui non è ancora stata individuata la decarbossilasi, con liberazione del C1 in forma di CO2.
Infine lo xilulosio può entrare nella via del pentoso fosfato a seguito della sua fosforilazione a xilulosio-5-fosfato nella reazione catalizzata dalla xilulochinasi o ATP:D-xilulosio-5-fosfotransferasi (EC 2.7.1.17).[23]
Galattosemia
La galattosemia è una malattia metabolica genetica causata da un difetto a carico di uno dei geni che codificano per gli enzimi della via di Leloir.
Descritta per la prima volta da von Reuss nel 1908, è caratterizzata da un anomalo accumulo di galattonato e galattitolo in molti organi del corpo.[5]
Tra i sintomi si ritrova la cataratta, che spesso compare precocemente, entro i primi due anni di vita, mentre nei casi più severi si hanno anche danni ai reni, al fegato e al cervello.
Riguardo alla cataratta sembra che tra i fattori scatenanti ci siano anche la sintesi e l’accumulo di galattitolo nel cristallino in quanto:
- essendo osmoticamente attivo, determina un aumento della pressione osmotica con conseguente richiamo d’acqua nella cellula e possibile rottura della membrana plasmatica;[19]
- la sua sintesi causa una deplezione di NADPH che, riducendo l’attività della glutatione reduttasi (EC 1.8.1.7), determina un incremento dei radicali liberi e quindi dello stress ossidativo.[24]
Attualmente, lo standard terapeutico per la cura della galattosemia è una dieta a ridotto contenuto di galattosio.[25]
Bibliografia
- ^ a b c Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 8th Edition. W.H. Freeman and Company, 2021.
- ^ a b c d Rosenthal M.D., Glew R.H. Medical biochemistry – Human metabolism in health and disease. John Wiley & sons, 2009.
- ^ a b Leloir L.F., de Fekete M.A., Cardini C.E. Starch and oligosaccharide synthesis from uridine diphosphate glucose. J Biol Chem 1961;236:636-41. doi:10.1016/S0021-9258(18)64280-2
- ^ a b c d e Conte F., van Buuringen N., Voermans N.C., Lefeber D.J. Galactose in human metabolism, glycosylation and congenital metabolic diseases: time for a closer look. Biochim Biophys Acta Gen Subj 2021;1865(8):129898. doi:10.1016/j.bbagen.2021.129898
- ^ a b Timson D.J. Type IV galactosemia. Genet Med 2019;21:1283-1285. doi:10.1038/s41436-018-0359-z
- ^ Pasteur L. Note sur le sucre de lait. 1856.
- ^ Berthelot M. Chimie organique fondée sur la synthèse Vol. 1, 1860. Mallet-Bachelier.
- ^ Luis Leloir – Biographical. NobelPrize.org. Nobel Prize Outreach 2026. Fri. 6 Feb 2026. https://www.nobelprize.org/prizes/chemistry/1970/leloir/biographical/
- ^ National Center for Biotechnology Information. PubChem Compound Summary for CID 6036, D-Galactose. https://pubchem.ncbi.nlm.nih.gov/compound/D-Galactose. Accessed Sept. 24, 2025.
- ^ a b Soderberg T. Organic chemistry with a biological emphasis. Volume II. Chemistry Publications. 2019. https://digitalcommons.morris.umn.edu/chem_facpubs/2
- ^ a b Tefsen B., Ram A.F.J., van Die I., Routier F.H. Galactofuranose in eukaryotes: aspects of biosynthesis and functional impact. Glycobiology 2012;22(4):456-469. doi:10.1093/glycob/cwr144
- ^ Flynn A. Galactosemia. Editor(s): McSweeney P.L.H., McNamara J.P. Encyclopedia of dairy sciences. 3rd Edition. Academic Press, 2022;853-858. doi:10.1016/B978-0-12-818766-1.00094-5
- ^ de la Rosa L.A., Alvarez-Parrilla E., Gonzàlez-Aguilar G.A. Fruit and vegetable phytochemicals: chemistry, nutritional value, and stability. 1st Edition. John Wiley & Sons, 2010.
- ^ Acosta P.B., Gross K.C. Hidden sources of galactose in the environment. Eur J Pediatr 1995;154(7 Suppl 2):S87-92. doi:10.1007/BF02143811
- ^ Gross K. C., Acosta P. B. Fruits and vegetables are a source of galactose: implications in planning the diets of patients with galactosaemia. J Inherit Metab Dis 1991;14(2):253-258. doi:10.1007/BF01800599
- ^ Zecca L., Mesonero J.E., Stutz A., Poirée J.C., Giudicelli J., Cursio R., Gloor S.M., Semenza G. Intestinal lactase-phlorizin hydrolase (LPH): the two catalytic sites; the role of the pancreas in pro-LPH maturation. FEBS Lett 1998;435(2-3):225-8. doi:10.1016/s0014-5793(98)01076-x
- ^ Richmond M.L., Gray J.I., and Stine C.M. Beta-galactosidase: review of recent research related to technological application, nutritional concerns, and immobilization. J Dairy Sci 1981;64:1759-1771. doi:10.3168/jds.S0022-0302(81)82764-6
- ^ a b Wright E.M., Hirayama B.A., Loo D.F. Active sugar transport in health and disease. J Intern Med 2007;261(1):32-43. doi:10.1111/j.1365-2796.2006.01746.x
- ^ a b c d e Coelho A.I., Rubio-Gozalbo M.E., Vicente J.B., Rivera I. Sweet and sour: an update on classic galactosemia. J Inherit Metab Dis 2017;40(3):325-342. doi:10.1007/s10545-017-0029-3
- ^ Thoden J.B., Timson D.J., Reece R.J., and M. Holden H.M. Molecular structure of human galactose mutarotase. J Biol Chem 2004;279(22):23431-23437. doi:10.1074/jbc.M402347200
- ^ a b Holden H.M., Rayment I., Thoden J.B. Structure and function of enzymes of the Leloir pathway for galactose metabolism. J Biol Chem 2003;278(45):43885-43888. doi:10.1074/jbc.r300025200
- ^ Leslie N., Yager C., Reynolds R., Segal S. UDP-galactose pyrophosphorylase in mice with galactose-1-phosphate uridyltransferase deficiency. Mol Genet Metab 2005;85(1):21-7. doi:10.1016/j.ymgme.2005.01.004
- ^ Lai K., Klapa M.I. Alternative pathways of galactose assimilation: could inverse metabolic engineering provide an alternative to galactosemic patients? Metab Eng 2004;6(3):239-44. doi:10.1016/j.ymben.2004.01.001
- ^ Pintor J. Sugars, the crystalline lens and the development of cataracts. Biochem Pharmacol 2012;1:4.
- ^ Coelho A.I., Berry G.T., Rubio-Gozalbo M.E. Galactose metabolism and health. Curr Opin Clin Nutr Metab Care 2015;18(4):422-427. doi:10.1097/MCO.0000000000000189
Domande Frequenti
Che tipo di zucchero è il galattosio e com'è fatto?
Il galattosio è un monosaccaride aldoesoso a sei atomi di carbonio, isomero del glucosio. In soluzione esiste in equilibrio termodinamico tra una forma instabile a catena aperta e quattro forme cicliche anomeriche, note come anomeri alfa e beta (piranosici e furanosici).
Come viene metabolizzato il galattosio nell'organismo?
Dopo l'assorbimento intestinale tramite il trasportatore SGLT1, il galattosio raggiunge il fegato. Qui viene convertito in glucosio principalmente attraverso la via di Leloir, una sequenza di quattro reazioni enzimatiche. Vie secondarie includono la via dei polioli e l'ossidazione.
Quali sono le cause e le conseguenze della galattosemia?
La galattosemia è una malattia genetica causata dal deficit di uno degli enzimi della via di Leloir. Questo blocco determina un accumulo tossico di galattitolo e galattonato nei tessuti, provocando danni a fegato, reni e sistema nervoso, oltre alla comparsa precoce di cataratta.