Amylopectin is a highly branched polysaccharide composed of α-D-glucose units. Together with amylose, it is one of the two main constituents of starch granules, the primary form of energy storage in plants and the most widespread and abundant form of carbohydrate reserve on Earth.[1]
Glucose monomers are connected by α-(1→4) glycosidic bonds to form linear chains, while branches are introduced through α-(1→6) glycosidic bonds.[2]
Its biosynthesis requires the coordinated activity of at least four distinct classes of enzymes: starch synthases (EC 2.4.1.21), starch branching enzymes (EC 2.4.1.18), starch debranching enzymes, and starch phosphorylase (EC 2.4.1.1).[3]
Within starch granules, amylopectin is more abundant than amylose and forms a semi-crystalline matrix in which amylose appears to be embedded.
The amylose/amylopectin ratio significantly affects the physicochemical properties of starch, thereby influencing both its industrial applications, such as the production of food additives, and its potential health effects.[4][5]
Summary: Key Points
- Starch matrix: amylopectin is a giant, highly branched polysaccharide that forms the primary semi-crystalline framework of plant starch granules.
- Chemical linkages: linear glucose chains are connected by α-(1→4) glycosidic bonds, while the characteristic branch points are introduced via α-(1→6) bonds.
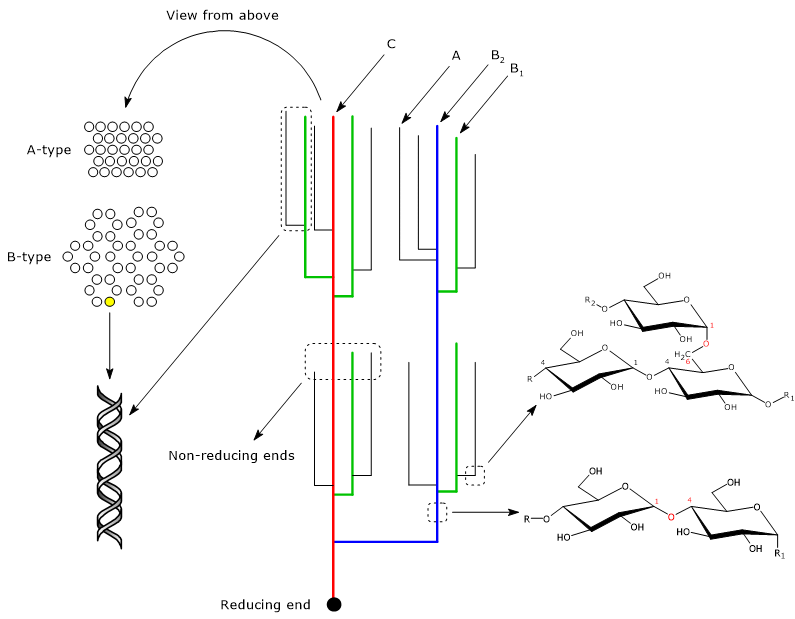
- Cluster architecture: chains are classified into outer (A), inner (B), and reducing-end (C) types, assembling into dense crystalline polymorphs (A-type in cereals) or open hydrated structures (B-type in tubers).
- Coordinated biosynthesis: its synthesis relies on the strict cooperation of starch synthases (SSI–SSV), starch branching enzymes (SBEs), debranching enzymes, and starch phosphorylase.
Contents
- Structure of amylopectin
- Amylopectin synthesis
- Phosphorylation of amylopectin
- Amylose/amylopectin ratio
- References
Structure of amylopectin
Amylopectin is a highly branched polysaccharide with a molecular weight on the order of 107-108 Daltons, making it much larger than amylose. It is composed of 104-105 glucose residues linked by α-(1→4) glycosidic bonds to form numerous relatively short chains, with a degree of polymerization ranging from 18 to 25 units.[6] The chain length varies depending on the source of starch as well as the environmental and nutritional conditions during plant growth and seed development.[2]
The chains are interconnected by α-(1→6) glycosidic bonds, giving rise to a tree-like architecture in which neighboring chains assemble into a cluster-like structure.[7]
In most starches, α-(1→6) linkages account for about 5% of the total glycosidic bonds, a lower proportion than in glycogen (≈9%), where the branches are more evenly distributed. The length and distribution of branches strongly influence the physicochemical properties of amylopectin, including solubility, viscosity, retrogradation behavior, and the gelatinization and pasting temperatures. For example, glycogen is water-soluble, whereas amylopectin and starch are not.[5]
Amylopectin chains
Amylopectin chains can be classified either by their length or by the presence or absence of branching.
Length-based classification
Two main types of chains are distinguished: short and long. Short chains have a degree of polymerization (DP) ranging from 6 to 36 glucosyl units, although the upper limit depends on the amylopectin source. Long chains are defined as those with a DP ≥ 36. In most starches, the molar ratio of long to short chains is approximately 19:6, and is generally higher in A-type crystalline starches (e.g., cereal endosperm starches) than in B-type crystalline starches (e.g., potato starch).[6]
Branching-based classification
According to their connections to other chains, amylopectin chains are divided into three categories: A-chains, B-chains, and C-chains.[8]
- A-chains are unbranched, short chains with an average DP of about 13; they represent the outer chains of the molecule.
- B-chains contain at least one branch (either an A- or another B-chain), are longer than A-chains, and are located in the inner regions of the molecule. They are further subclassified into B1-chains (DP ≈ 22), B2-chains (DP ≈ 42), B3-chains (DP ≈ 69), B4-chains, and so forth.
- The C-chain is a special type of B-chain that carries the sole reducing end of the amylopectin molecule.
A-chains and B1-chains participate in the formation of clusters, while B2-, B3-, and B4-chains are thought to span two, three, and four clusters, respectively.[8][9]
| Classification criteria | Chain type | Degree of polymerization (DP) | Structural position and function |
|---|---|---|---|
| Length-based | Short chains | 6 to 36 glucosyl units | Predominant in A-type crystalline starches (e.g., cereal endosperm) |
| Long chains | ≥ 36 glucosyl units | More frequent in B-type crystalline starches (e.g., potato starch) | |
| Branching-based | A-chains | Average DP ≈ 13 | Unbranched outer chains; participate in cluster formation |
| B-chains | DP varies (B1 ≈ 22, B2 ≈ 42, B3 ≈ 69) | Branched inner chains; span across one or multiple clusters | |
| C-chain | Variable (Special B-chain) | Carries the sole reducing end of the entire amylopectin molecule |
A-type, B-type and C-type polymorphs
Within the clusters, neighboring linear chain segments form double helices that run parallel to each other, with a periodicity of 2.1 nm, where each turn comprises six glucose residues per chain.[10] These double helices assemble into two distinct crystalline structures: the A-type polymorph, which is denser and typical of cereal grains, and the B-type polymorph, which has a more open hexagonal structure, is less dense and more hydrated, and is characteristic of tuber and high-amylose starches. The C-type polymorph represents a mixture of A- and B-type structures and is found in the starches of roots, legumes, and certain fruits.
This organization underlies the semi-crystalline nature of starch granules.[2]
| Polymorph | Structure and density | Hydration level | Typical botanical sources |
|---|---|---|---|
| A-type | Dense, parallel double helices | Low hydration | Cereal grains (e.g., cereal endosperm starches) |
| B-type | Open hexagonal structure, less dense | Highly hydrated | Tubers (e.g., potato starch) and high-amylose starches |
| C-type | Mixture of A-type and B-type structures | Variable / Intermediate | Roots, legumes, and certain fruits |
Growth rings
Although starch granules vary in shape, their internal architecture is remarkably conserved across species. When observed under a microscope, most starches display a regular pattern of alternating light and dark rings, known as growth rings because of their resemblance to tree rings.[10]
The growth rings surround the hilum, the central region of the granule, whose precise structure remains unclear, although it appears to consist of a relatively disordered α-glucan arrangement. Each ring has a thickness of 200–400 nm and reflects the alternation of less dense amorphous regions and denser semi-crystalline regions.[10]
According to the cluster model, the semi-crystalline regions arise from the alternation of crystalline and amorphous lamellae, stacked with a periodicity of about 9–10 nm.[9] The crystalline lamellae, composed of amylopectin linear chains arranged in A-, B-, or C-type polymorphs, extend about 6–7 nm, whereas the amorphous lamellae, which contain most of the branch points, extend about 3 nm.[11][12]
Amylopectin synthesis
Amylopectin synthesis is thought to initiate at the hilum of the starch granule.[13]
It requires the coordinated activities of at least four distinct classes of enzymes: starch synthases, starch phosphorylase, starch branching enzymes, and starch debranching enzymes.[1] Each class comprises several isoforms with distinct biochemical properties.
Like amylose biosynthesis, amylopectin synthesis depends on short malto-oligosaccharides (MOS), α-(1→4)-glucans with a DP of 2 to 7, which act as primers and are elongated by starch synthase, in a manner similar to the role of glycogenin during the early stages of glycogen synthesis.[14]
MOS appear to originate from multiple sources, all of which require the action of enzymes involved in starch biosynthesis, including:
- starch synthase III (SSIII) and starch phosphorylase (the latter in cooperation with the disproportionating enzyme, EC 2.4.1.25), which use ADP-glucose and glucose-1-phosphate as substrates, respectively;
- starch debranching enzymes, which generate MOS during the trimming of other amylopectin molecules.[14]
Because MOS exhibit low solubility in aqueous environments, they seem to evade hydrolysis by alpha-amylase (EC 3.2.1.1) and beta-amylase (EC 3.2.1.2).[6]
The spatial and temporal coordination of the enzymes involved, many of which physically interact to form multienzyme complexes, is essential for converting the products of photosynthesis into the highly organized and insoluble polysaccharide structure.[5] Moreover, as in amylose and glycogen synthesis, the polymerization of glucose into amylopectin, and more generally of osmotically active monosaccharides into osmotically inert polysaccharides, enables the storage of large amounts of carbohydrates within the cell without causing a substantial increase in osmotic pressure.[15]
Starch synthase
Six isoforms of starch synthase have been identified, all structurally related. Five of these, starch synthase I, II, III, IV, and V (SSI, SSII, SSIII, SSIV, and SSV; EC 2.4.1.21), are involved in amylopectin biosynthesis. They are localized either in the plastid stroma or partitioned between the stroma and starch granules. The sixth isoform, granule-bound starch synthase (GBSS; EC 2.4.1.242), is almost exclusively associated with granules and is responsible for amylose synthesis.[10][16]
The first four isoforms are catalytically active and, like GBSS, glycogen phosphorylase (EC 2.4.1.1), and glycogen synthase (EC 2.4.1.11), which are enzymes involved in glycogenolysis and glycogen synthesis, belong to the glycosyltransferase family (EC 2.4). In contrast, starch synthase V lacks catalytic activity.
During amylopectin synthesis, starch synthase catalyzes the addition of one glucose residue to the non-reducing end of a pre-existing α-(1→4)-linked glucan chain, forming a new α-(1→4) glycosidic bond.[12]
(1→4)-α-D-glucosyl(n) + ADP-α-D-glucose ⇌ (1→4)-α-D-glucosyl(n+1) + ADP + H+
Unlike glycogen synthase, which uses UDP-glucose, starch synthase employs ADP-glucose as the glucosyl donor.[5]
The mode of action of starch synthases I–IV differs from that of GBSS. While SS isoforms catalyze the incorporation of only one glucose unit per substrate encounter, a distributive mode of action, GBSS is capable of adding multiple glucose units per encounter, a processive mode of action.[6]
Roles of starch synthases
The initial steps of amylopectin biosynthesis, as well as the formation of normal starch granules, require the presence of SSIV, although SSIII also appears to contribute, with partially overlapping functions.[14]
Like GBSS, SSIV depends on a member of the PTST protein family, PTST2, which lacks catalytic activity but facilitates enzyme binding to α-glucans through a specific carbohydrate-binding domain. SSIV can also dimerize, a feature important both for its catalytic activity and for protein–protein interactions.
According to a proposed model of PTST2 function, the protein recognizes malto-oligosaccharides with a specific three-dimensional helical conformation via its carbohydrate-binding domain. PTST2 then forms a complex with MOS, which subsequently interacts with an SSIV dimer. The dimer catalyzes the elongation of the α-glucan, while PTST2 is released, enabling it to bind another MOS and mediate further interactions with SSIV dimers.[17]
Following the action of SSIV, the other starch synthase isoforms extend the glucan chains. SSI elongates malto-oligosaccharides with a DP of 6–7 to form oligosaccharides with a DP of 8–12. These, in turn, are excellent substrates for SSII, which extends them to chains with a DP of 12–30. SSIII further elongates the glucans, producing linear chains with DP values greater than 30. Thus, SSIII appears to function not only in the early stages of granule initiation but also during later phases of starch biosynthesis.[1]
Finally, SSIV and SSV seem to be required for the production of a regular number of starch granules with normal morphology.[7][14]
Starch branching enzymes
Starch branching enzymes (SBEs) catalyze the formation of α-(1→6) glycosidic bonds, thereby introducing branch points into linear α-glucan chains, such as those in glycogen and amylopectin.[18][19] Their action increases the number of non-reducing ends, which serve as acceptors of glucose units during elongation reactions.[20]
SBEs function by cleaving an internal α-(1→4) bond within an α-glucan chain, releasing an oligosaccharide (or oligonucleotide) whose reducing end is subsequently attached to the hydroxyl group at the C6 position of a glucosyl residue on an α-glucan chain. The result is a new α-(1→6) linkage. The acceptor chain can be either the same chain from which the oligosaccharide was removed (intra-chain transfer) or a different chain (inter-chain transfer).
One of the factors influencing the type of transfer appears to be the relative concentration of linear α-(1→4) chains. In particular, closely associated chains, such as those forming double helices within clusters, tend to favor inter-chain transfer.[21] Moreover, the interaction between starch synthase I and starch branching enzymes seems to play a key role in determining the bimodal chain-length distribution characteristic of plant starches.[14]
Starch branching enzyme isoforms
Plants possess two isoforms of starch branching enzyme, referred to as SBEI and SBEII. Encoded by different genes, they exhibit distinct biochemical properties, suggesting that they play different roles in determining the structure of amylopectin and amylose.[6]
SBEI is expressed predominantly in storage tissues, indicating an important role in defining the structural properties of storage starches. It shows a substrate preference for amylose and can transfer oligosaccharides with a DP greater than 30, although most transfers involve chains of DP 10–13. SBEI also appears to be involved in the synthesis of super-long (or extra-long) amylopectin chains, while its other contributions to amylopectin structure seem less significant. Not all plants express SBEI; for example, Arabidopsis and canola (Brassica napus L.), both oil-storing plants, possess only SBEII, and starch is found exclusively in their photosynthetic tissues.[22][23]
SBEII is widely expressed in grasses, cereals, and many other plants. Loss of SBEII activity results in pronounced alterations to amylopectin architecture and a reduction in starch content. The enzyme has a substrate preference for amylopectin and transfers oligosaccharides with a DP of 6–14. In cereals and grasses, two tissue-specific isoforms are encoded by distinct genes: SBEIIa, expressed mainly in leaves, and SBEIIb, expressed mainly in the endosperm.[6]
Starch debranching enzymes
Starch debranching enzymes catalyze the hydrolysis of α-(1→6) glycosidic bonds and are members of the α-amylase superfamily.
Two types of debranching enzymes are present in plants: isoamylases (EC 3.2.1.68) and pullulanases (EC 3.2.1.41).[24] Isoamylases act on amylopectin and other polyglucans, whereas pullulanases debranch both amylopectin and pullulan, a fungal polysaccharide. Isoamylases and pullulanases also differ in substrate specificity: isoamylases hydrolyze branches composed of at least three glycosidic residues, while pullulanases can act on branches of just two.[25]
During starch granule formation, starch debranching enzymes play a crucial role in defining the water-insoluble properties and fine structure of amylopectin. Enzyme activity is thought to promote clustering of the remaining branches, thereby enhancing interactions between adjacent chains and facilitating α-helix formation. This process, in turn, appears essential for the development of the semi-crystalline structures of amylopectin and, consequently, of starch. In these semi-crystalline regions, branches are presumably inaccessible to starch debranching enzymes, α-amylase, and β-amylase.[6]
Starch debranching enzymes are also employed industrially, particularly in the production of resistant starch and cyclodextrins, which are cyclic oligosaccharides.[25]
Starch phosphorylase
Like starch synthases, starch phosphorylase belongs to the family of glycosyltransferases and closely resembles glycogen phosphorylase. In plants, it occurs in at least two isoenzymatic forms: Pho1, located in the stroma of plastids and considered the true starch phosphorylase involved in starch synthesis, and Pho2, which is cytosolic.[26]
Starch phosphorylase is thought to participate in the initial steps of starch synthesis by catalyzing the reversible transfer of glucosyl units to an α-glucan, where they are linked by an α-(1→4) glycosidic bond.[5] Unlike starch synthases, starch phosphorylase uses glucose-1-phosphate, not ADP-glucose, as the glucosyl donor.[26]
Phosphorylation of amylopectin
Amylopectin, like glycogen, carries phosphate groups in variable amounts depending on the botanical source of the starch. For example, potato starch contains relatively high levels of phosphate groups, with a degree of substitution of about 0.1%–0.3%, whereas cereal endosperm starches generally have phosphate contents below 0.01%.[27]
Phosphorylation of amylopectin is catalyzed by two dikinases present in plastids: α-glucan water dikinase (EC 2.7.9.4) and phospho-glucan water dikinase (EC 2.7.9.5). These enzymes transfer the β-phosphate group of ATP to a glucosyl unit of an α-glucan chain, while the γ-phosphate group is transferred to water. Specifically, α-glucan water dikinase phosphorylates the hydroxyl group at the C6 position, whereas phospho-glucan water dikinase phosphorylates the hydroxyl group at the C3 position, usually on a prephosphorylated glucan chain. About two-thirds of phosphate groups are bound at the C6 position and 20%–30% at the C3 position. Phosphate groups are also detected at the C2 position, but in much smaller amounts. The enzyme responsible for this phosphorylation is not known.[28][29]
With regard to substrate specificity, phosphorylation appears to accumulate more readily on longer chains. Furthermore, there seems to be an inverse correlation between total phosphate content and the frequency of amylopectin branching. The negative charges carried by phosphate groups cause mutual repulsion between neighboring phosphorylated oligosaccharides. These repulsions seem to promote chain opening and hydration, thereby influencing the activity of biosynthetic enzymes and making the chains more susceptible to attack by amylases.[30][31]
Amylose/amylopectin ratio
Starch granules are composed primarily of amylopectin and amylose.[1] The relative proportions of the two polysaccharides vary, with amylose typically accounting for no more than 35% of the granule’s dry weight.[12] However, some plants produce starch granules composed mostly, or almost entirely, of amylopectin, (known as waxy starches), while others produce starch granules composed predominantly, or almost exclusively, of amylose.[21]
The amylose/amylopectin ratio strongly influences the physicochemical properties of starch, including water absorption, gelatinization, retrogradation, and resistance to enzymatic hydrolysis. The latter property is especially relevant during carbohydrate digestion, as it determines the rate at which amylose and amylopectin are hydrolyzed by α-amylase to maltose and maltotriose. Consequently, the amylose/amylopectin ratio affects both the health impacts of different starch types and their industrial applications.[5][32][33]
References
- ^ a b c d Qu J., Xu S., Zhang Z., Chen G., Zhong Y., Liu L., Zhang R., Xue J., Guo D. Evolutionary, structural and expression analysis of core genes involved in starch synthesis. Sci Rep 2018;8(1):12736. doi:10.1038/s41598-018-30411-y
- ^ a b c Cornejo-Ramírez Y.I., Martínez-Cruz O., Del Toro-Sánchez C.L., Wong-Corral F.J., Borboa-Flores J. & Cinco-Moroyoqui F.J. The structural characteristics of starches and their functional properties. CYTA J Food 2018;16(1):1003-1017. doi:10.1080/19476337.2018.1518343
- ^ Smith A.M., Zeeman S.C. Starch: a flexible, adaptable carbon store coupled to plant growth. Annu Rev Plant Biol 2020;71:217-245. doi:10.1146/annurev-arplant-050718-100241
- ^ Junejo S.A., Flanagan B.M., Zhang B., Dhital S. Starch structure and nutritional functionality – Past revelations and future prospects. Carbohydr Polym 2022;277:118837. doi:10.1016/j.carbpol.2021.118837
- ^ a b c d e f Crofts N., Abe N., Oitome N.F., Matsushima R., Hayashi M., Tetlow I.J., Emes M.J., Nakamura Y., Fujita N. Amylopectin biosynthetic enzymes from developing rice seed form enzymatically active protein complexes. J Exp Bot 2015;66(15):4469-82. doi:10.1093/jxb/erv212
- ^ a b c d e f g Tetlow I.J., Bertoft E. A review of starch biosynthesis in relation to the building block-backbone model. Int J Mol Sci 2020;21(19):7011. doi:10.3390/ijms21197011
- ^ a b Abt M.R., Pfister B., Sharma M., Eicke S., Bürgy L., Neale I., Seung D., Zeeman S.C. STARCH SYNTHASE5, a noncanonical starch synthase-like protein, promotes starch granule initiation in Arabidopsis. Plant Cell 2020;32(8):2543-2565. doi:10.1105/tpc.19.00946
- ^ a b Li G., Yacine Y., Zhu F. Relationships between supramolecular organization and amylopectin fine structure of quinoa starch. Food Hydrocoll 2021;117:106685. doi:10.1016/j.foodhyd.2021.106685
- ^ a b Pfister B., Zeeman S.C., Rugen M.D., Field R.A., Ebenhöh O., Raguin A. Theoretical and experimental approaches to understand the biosynthesis of starch granules in a physiological context. Photosynth Res 2020;145:55-70. doi:10.1007/s11120-019-00704-y
- ^ a b c d Pfister B., Zeeman S.C. Formation of starch in plant cells. Cell Mol Life Sci 2016;73(14):2781-807. doi:10.1007/s00018-016-2250-x
- ^ Jenkins P.J., Donald A.M. The influence of amylose on starch granule structure. Int J Biol Macromol 1995;17(6):315-21. doi:10.1016/0141-8130(96)81838-1
- ^ a b c Gous P.W., Fox G.P. Review: Amylopectin synthesis and hydrolysis – Understanding isoamylase and limit dextrinase and their impact on starch structure on barley (Hordeum vulgare) quality. Trends Food Sci Technol 2017;62:23-32. doi:10.1016/j.tifs.2016.11.013
- ^ Ziegler G.R., Creek J.A., Runt J. Spherulitic crystallization in starch as a model for starch granule initiation. Biomacromolecules 2005;6(3):1547-54. doi:10.1021/bm049214p
- ^ a b c d e Szydlowski N., Ragel P., Raynaud S., Lucas M.M., Roldán I., Montero M., Muñoz F.J., Ovecka M., Bahaji A., Planchot V., Pozueta-Romero J., D’Hulst C., Mérida A. Starch granule initiation in Arabidopsis requires the presence of either class IV or class III starch synthases. Plant Cell 2009;21(8):2443-57. doi:10.1105/tpc.109.066522
- ^ Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 8th Edition. W.H. Freeman and Company, 2021.
- ^ Zeeman S.C., Kossmann J., Smith A.M. Starch: its metabolism, evolution, and biotechnological modification in plants. Annu Rev Plant Biol 2010;61:209-34. doi:10.1146/annurev-arplant-042809-112301
- ^ Seung D., Boudet J., Monroe J., Schreier T.B., David L.C., Abt M., Lu K.J., Zanella M., Zeeman S.C. Homologs of PROTEIN TARGETING TO STARCH control starch granule initiation in Arabidopsis leaves. Plant Cell 2017;29(7):1657-1677. doi:10.1105/tpc.17.00222
- ^ Ball S.G., Morell M.K. From bacterial glycogen to starch: understanding the biogenesis of the plant starch granule. Annu Rev Plant Biol. 2003;54:207-33. doi:10.1146/annurev.arplant.54.031902.134927
- ^ Wilkens C., Svensson B., Møller M.S. Functional roles of starch binding domains and surface binding sites in enzymes involved in starch biosynthesis. Front Plant Sci 2018;9:1652. doi:10.3389/fpls.2018.01652
- ^ Sawada T., Nakamura Y., Ohdan T., Saitoh A., Francisco P.B. Jr, Suzuki E., Fujita N., Shimonaga T., Fujiwara S., Tsuzuki M., Colleoni C., Ball S. Diversity of reaction characteristics of glucan branching enzymes and the fine structure of α-glucan from various sources. Arch Biochem Biophys 2014;562:9-21. doi:10.1016/j.abb.2014.07.032
- ^ a b Wang J., Hu P., Lin L., Chen Z., Liu Q., Wei C. Gradually decreasing starch branching enzyme expression is responsible for the formation of heterogeneous starch granules. Plant Physiol 2018;176(1):582-595. doi:10.1104/pp.17.01013
- ^ Guan H.P., Preiss J. Differentiation of the properties of the branching isozymes from maize (Zea mays). Plant Physiol 1993;102(4):1269-1273. doi:10.1104/pp.102.4.1269
- ^ Dumez S., Wattebled F., Dauvillee D., Delvalle D., Planchot V., Ball S.G., D’Hulst C. Mutants of Arabidopsis lacking starch branching enzyme II substitute plastidial starch synthesis by cytoplasmic maltose accumulation. Plant Cell 2006;18(10):2694-709. doi:10.1105/tpc.105.037671
- ^ Møller M.S., Henriksen A., Svensson B. Structure and function of α-glucan debranching enzymes. Cell Mol Life Sci 2016;73(14):2619-41. doi:10.1007/s00018-016-2241-y
- ^ a b Xia W., Zhang K., Su L., Wu J. Microbial starch debranching enzymes: developments and applications. Biotechnol Adv 2021;50(3):107786. doi:10.1016/j.biotechadv.2021.107786
- ^ a b Cuesta-Seijo J.A., Ruzanski C., Krucewicz K., Meier S., Hägglund P., Svensson B., Palcic M.M. Functional and structural characterization of plastidic starch phosphorylase during barley endosperm development. PLoS One 2017;12(4):e0175488. doi:10.1371/journal.pone
- ^ Nitschke F., Wang P., Schmieder P., Girard J.M., Awrey D.E., Wang T., Israelian J., Zhao X., Turnbull J., Heydenreich M., Kleinpeter E., Steup M., Minassian B.A. Hyperphosphorylation of glucosyl C6 carbons and altered structure of glycogen in the neurodegenerative epilepsy Lafora disease. Cell Metab 2013;17(5):756-67. doi:10.1016/j.cmet.2013.04.006
- ^ Hizukuri S., Tabata S., Kagoshima Nikuni Z. Studies on starch phosphate Part 1. Estimation of glucose-6-phosphate residues in starch and the presence of other bound phosphate(s). Starch/Stärke 1970;22:338-343. doi:https://doi.org/10.1002/star.19700221004
- ^ Ritte G., Heydenreich M., Mahlow S., Haebel S., Kötting O., Steup M. Phosphorylation of C6- and C3-positions of glucosyl residues in starch is catalysed by distinct dikinases. FEBS Lett 2006;580(20):4872-6. doi:10.1016/j.febslet.2006.07.085
- ^ Zhou W., He S., Naconsie M., Ma Q., Zeeman S.C., Gruissem W. & Zhang P. Alpha-glucan, water dikinase 1 affects starch metabolism and storage root growth in Cassava (Manihot esculenta Crantz). Sci Rep 2017;7:9863 doi:10.1038/s41598-017-10594-6
- ^ Baunsgaard L., Lütken H., Mikkelsen R., Glaring M.A., Pham T.T., Blennow A. A novel isoform of glucan, water dikinase phosphorylates pre-phosphorylated alpha-glucans and is involved in starch degradation in Arabidopsis. Plant J 2005;41(4):595-605. doi:10.1111/j.1365-313X.2004.02322.x
- ^ Tester R.F., Morrison W.R. Swelling and gelatinization of cereal starches. I. Effects of amylopectin, amylose and lipids. Cereal Chem 1990;67:551-557.
- ^ Hoover R. Composition, molecular structure, and physicochemical properties of tuber and root starches: a review. Carbohydr Polym 2001;45(3):253-267 doi:10.1016/S0144-8617(00)00260-5
Domande Frequenti
What is the difference between amylopectin and amylose?
Amylopectin is a large, highly branched polysaccharide with alpha-(1→4) and alpha-(1→6) glycosidic bonds, making up the major crystalline matrix of starch. In contrast, amylose is smaller, predominantly linear with alpha-(1→4) bonds, and sits embedded within the amylopectin matrix.
What are the structural differences between A, B, and C-type starch polymorphs?
They are crystalline structures formed by amylopectin double helices. A-type is dense and common in cereal grains; B-type is a more open, highly hydrated hexagonal structure typical of tubers; C-type is a natural mixture of both A and B types found in roots and legumes.
Which enzymes are required for amylopectin biosynthesis?
Amylopectin synthesis requires the coordinated activity of four enzyme classes: starch synthases (SSI to SSV) for chain elongation, starch branching enzymes (SBEI and SBEII) for introducing branch points, starch debranching enzymes for structural trimming, and starch phosphorylase.