Lipid digestion and absorption are complex processes. They involve soluble enzymes and substrates with different degrees of solubility, occurring primarily in the stomach and small intestine.
Dietary lipids include triglycerides, phospholipids, steroids (especially cholesterol and its esters), fat-soluble vitamins (A, D, E, and K), and carotenoids.
Despite scientific societies recommending a lipid intake, basically triacylglycerols, not exceeding 30% of the daily caloric intake, they provide between 30% and 45% of total calories in the Western diet.
The distinctive hydrophobicity of many dietary lipids, while making triglycerides excellent molecules for energy storage, creates obstacles during digestion, intestinal absorption, and subsequent circulation. Indeed, lipids such as long-chain triglycerides, cholesterol, and fat-soluble vitamin esters aggregate into large droplets within the gastrointestinal lumen. These droplets must then be emulsified to allow hydrolases to catalyze their breakdown, namely, lipid digestion occurs.
Summary: Key Points
- Dietary composition: triglycerides make up about 90% of dietary lipids, providing high energy density (9 kcal/g), alongside phospholipids, cholesterol, and fat-soluble vitamins.
- Sequential breakdown: lipid digestion begins in the mouth (lingual lipase), continues in the stomach (gastric lipase), and concludes in the small intestine via pancreatic enzymes.
- Emulsification: amphipathic molecules, including bile salts and surfactants like free fatty acids and phospholipids, reduce lipid droplet size to maximize enzyme surface area.
- Enzymatic specificity: acid lipases preferentially cleave short- and medium-chain fatty acids at the sn-3 position, while pancreatic lipase targets long-chain triglycerides at sn-1 and sn-3 with the aid of colipase.
- Broad-spectrum hydrolases: pancreatic juice also provides cholesterol esterase and phospholipaseA2 to break down sterol esters and phospholipids within bile salt micelles.
Contents
Dietary lipids: classification and features
Different types of lipids possess distinct chemical and physical properties that directly influence how they are processed in the intestinal tract for digestion and subsequent absorption.
- Triglycerides (triacylglycerols): they account for about 90% of dietary lipids and consist of one glycerol molecule esterified to three fatty acids, mostly long-chain fatty acids (16–20 carbon atoms). They have an energy density more than twice that of carbohydrates and proteins (about 9 kcal/g vs. about 4 kcal/g) and amino acids. They must release their fatty acids to be used as a source of energy.
- Phospholipids: the main constituents of biological membranes, they consist of one glycerol molecule esterified with two fatty acids at the sn-1 and sn-2 positions, and a phosphoric acid at the sn-3 position. In turn, the phosphate group binds a hydrophilic group, such as choline, serine, or inositol, via an ester bond. While the daily dietary intake is low (1–2 g), about 10–20 g of biliary phospholipids (mostly phosphatidylcholine) also pour into the small intestine daily.
- Cholesterol and steroids: they are components of biological membranes found exclusively in animal products, unlike triglycerides and plant sterols. In the small intestine, dietary cholesterol (which should not exceed 300 mg per day) mixes with about 1 g per day of biliary cholesterol. Both forms are mostly non-esterified (85–90%), which is the only form capable of being absorbed. Like vitamins, this steroid molecule does not serve as an energy source.
Additionally, dietary steroids include plant stanols and sterols, such as beta-sitosterol, which is not absorbed under physiological conditions.
Lipid digestion: steps, enzymes, and products
Lipid digestion begins in the mouth, continues in the stomach, and ends in the small intestine.
Enzymes involved in triacylglycerol digestion are called lipases (EC 3.1.1.3).
They are proteins that catalyze the partial hydrolysis of triglycerides into a mixture of free fatty acids and acylglycerols. There are several lipases, the most important of which is produced by the exocrine pancreas. The others are lingual lipase, gastric lipase, and breast milk lipase.
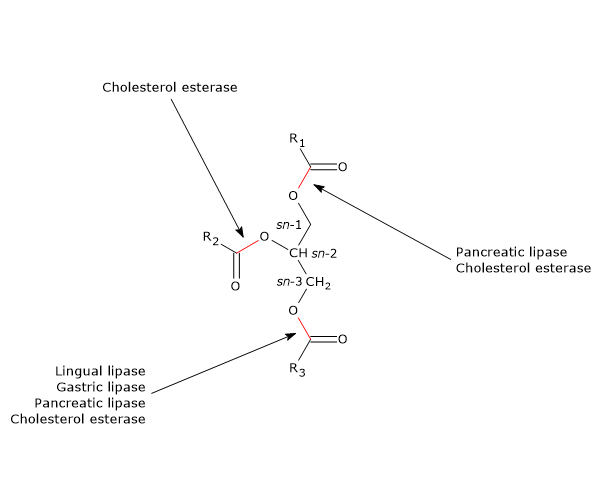
Other enzymes involved in lipid digestion are cholesterol esterase and phospholipases A1 and A2.
| Enzyme | Source/Location | Optimal pH | Target bond/Substrate | Primary products |
|---|---|---|---|---|
| Lingual lipase | Serous lingual glands/mouth & stomach | Acid-stable | sn-3 ester bonds of short/medium-chain triglycerides | 1,2-Diacylglycerols and 1 free fatty acid |
| Gastric lipase | Chief cells/stomach & upper duodenum | ≈4.0 (active 6.0–6.5) | sn-3 ester bonds of short/medium-chain triglycerides | 1,2-Diacylglycerols and 1 free fatty acid |
| Pancreatic lipase | Exocrine pancreas/jejunum (with colipase) | 7.0–8.8 | sn-1 and sn-3 ester bonds of long-chain triglycerides | 2-Monoacylglycerols and 2 free fatty acids |
| Breast milk lipase | Secreted in milk/small intestine | Neutral | Triglycerides (stimulated by bile salts in infant gut) | Free fatty acids and glycerol |
| Cholesterol esterase | Exocrine pancreas/bile salt micelles | Alkaline (stimulated by bile salts) | Broad: cholesterol esters, triglycerides, phospholipids, vit. A/D esters | Free cholesterol and fatty acids or glycerol |
| Phospholipase A2 | Exocrine pancreas (as zymogen)/micelles | Alkaline | sn-2 ester bonds of phospholipids | Lysophospholipids and 1 free fatty acid |
| Phospholipase A1 | Pancreatic juice/intestinal lumen | Alkaline | sn-1 ester bonds of phospholipids | Lysophospholipids and 1 free fatty acid |
Lingual lipase
In the mouth, food is broken into small particles and mixed with lingual lipase, an enzyme produced and secreted by serous lingual glands (von Ebner’s glands). This lipase is stable in an acid environment, allowing it to remain active in the stomach and even in the small intestine if proper pancreatic bicarbonate secretion is lacking.
The reaction catalyzed by lingual lipase releases a 1,2-diacylglycerol and a single free fatty acid, preferring short-chain fatty acids, such as butyric acid, or medium-chain fatty acids, such as caproic, caprylic, capric, and lauric acids. Notably, short-chain fatty acids are mainly esterified at the sn-3 position of the triacylglycerol.
However, lingual lipase plays a modest role in adult triacylglycerol digestion because:
- it has a significantly lower activity compared to pancreatic lipase;
- it acts primarily in the stomach, an environment where lipids tend to coalesce and form a separate phase, limiting the enzyme’s access to its substrates.
In the stomach, lipids are mixed also with gastric juice and hydrochloric acid. The acid also denatures the proteins of the lipid-protein complexes; the polypeptides are then digested by gastric proteases, and the lipids are released.
Physiological importance and role in emulsification
While minor in healthy adults, lingual lipase activity is critical for infants, whose pancreatic lipase is still immature. Infant nutrition benefits from this enzyme because milk triglycerides are rich in short- and medium-chain fatty acids. Furthermore, unlike pancreatic lipase, both lingual and gastric lipases can penetrate the fat globules of milk to initiate digestion. This pathway also becomes essential in adults during pancreatic insufficiency.
Beyond its role in lipid digestion, lingual lipase influences food choices. Because the tongue is sensitive to the taste of free fatty acids, especially polyunsaturated ones, rather than intact triglycerides, this enzymatic activity helps detect fatty foods as energy sources.
Finally, the release of short-chain fatty acids, medium-chain fatty acids, and diacylglycerols is crucial for lipid physics. Being amphipathic surfactants, these molecules orient their hydrophilic regions toward the aqueous phase and their hydrophobic regions toward the core of lipid droplets. This surface modification, combined with gastric churning, prevents droplets from coalescing and forms a stable emulsion. This finely dispersed emulsion is then released into the duodenum as chyme, providing a massive surface area for subsequent pancreatic hydrolases.
Gastric lipase
In the stomach, lipids are hydrolyzed also by a second acid lipase, the gastric lipase.
This enzyme is secreted by the chief cells of the gastric mucosa.
It has an optimal pH around 4, but is still quite active at less acidic pH values, 6 to 6.5. Therefore, it probably remains active even in the upper duodenum, where the pH is between 6 and 7.
The enzyme preferentially catalyzes the hydrolysis of triglycerides with short-chain fatty acids and medium-chain fatty acids, but may also hydrolyze long-chain fatty acids. Regardless of the type of fatty acids, gastric lipase preferentially cleaves those at the sn-3 position, leading to the release of a free fatty acid and a 1,2-diacylglycerol, molecules that can act as surfactants, as previously seen.
Like lingual lipase, it is particularly active on milk triglycerides, including those in breast milk, which are rich in short-chain and medium-chain fatty acids. Moreover, it is also active on the triacylglycerols of some tropical oils, e.g. coconut oil, which are rich in medium-chain fatty acids.
The enzyme can account for 10% to 30% of triacylglycerol hydrolysis occurring in the gastrointestinal tract, and up to 50% in breast-fed infants.
Bile salts and the emulsion of lipid droplets
The chyme, containing a lipid emulsion made up of droplets of diameter less than 0.5 mm, enters the upper portion of the small intestine, the duodenum, where lipid digestion, and the hydrolysis of triglycerides, continues.
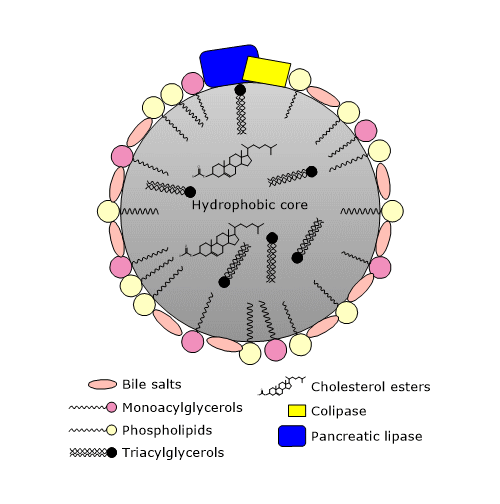
In the duodenum, the chyme is mixed with bile, whose release by the gallbladder is stimulated by cholecystokinin, a hormone secreted by cells of the mucosa of the duodenum and jejunum in response to the ingestion of a meal, particularly if high in fat.
In the bile, among the other components, there are bile salts, phospholipids, and cholesterol. Bile salts are bile acids conjugated with glycine or taurine. In turn, bile acids are oxygenated derivatives of cholesterol. Bile acids and bile salts are both synthesized by the liver. They are amphipathic molecules, in whose planar ring structure you can identify a hydrophobic face and a hydrophilic face. Therefore, they are able to further emulsify lipid droplets, increasing the surface area for hydrolytic enzyme activity.
In particular, salts of cholic acid, which contain three hydroxyl groups, are better emulsifiers than salts of deoxycholic acid, which instead contain only two hydroxyl groups.
Note: the gallbladder secretes about 30 g of bile salts each day, together with phospholipids and cholesterol. Most of the bile salts and cholesterol are then reabsorbed, so that the daily fecal loss of bile salts and steroids is quite low, 0.2–1 g.
The mechanism of peristalsis and the surfactants seen so far, that is, free fatty acids, acylglycerols, phospholipids, and bile salts, ensure the formation of microscopic micelles, which further increase the available surface areas for hydrolytic enzyme activity.
It should be underlined that triacylglycerols with short-chain and medium-chain fatty acids can be both hydrolyzed and absorbed in the absence of bile salts, although their presence increases the absorption.
Pancreatic lipase
Cholecystokinin also stimulates the exocrine pancreas to secrete a pancreatic juice containing, among other molecules, pancreatic lipase.
The enzyme catalyzes the digestion of the majority of ingested triglycerides, mainly in the upper portion of the jejunum.
It has an optimum pH of 7.0 to 8.8. Therefore it is not an acid lipase, unlike lingual and gastric lipases.
Pancreatic lipase catalyzes the cleavage of fatty acids, typically with 10 or more carbon atoms, primarily in sn-1 and sn-3 positions of the glycerol backbone. The products of the reaction are free fatty acids and 2-monoacylglycerols.
2-Monoacylglycerol, the primary form in which the monoacylglycerols are absorbed from the small intestine, can undergo an isomerization process in which the remaining fatty acid shifts to carbon 1 or 3. However, the rate of isomerization is slower than the rate of uptake of the molecule from the small intestine.
In vitro, pancreatic lipase is inhibited by bile salts, whereas in vivo, it hydrolyzes triglycerides in a very efficient manner, due to the presence of a protein cofactor secreted by the exocrine pancreas, the colipase. This protein has no catalytic activity, is produced in an inactive form, called procolipase, and is activated by trypsin in the duodenum. Lipid droplets are coated with phospholipids and bile salts, that give them a negative charge which prevents the binding of lipase, but attracts the colipase. In turn, colipase binds pancreatic lipase (lipase and colipase bind in a 1:1 molar ratio), thus anchoring the enzyme to the water-lipid interface of the lipid droplets.
Lipase in breast milk
This lipase has a neutral pH optimum, and is stimulated by bile salts. The enzyme contributes substantially to hydrolysis of the triglycerides in the intestine of breast-fed infants.
Instead, there are no lipases in cow’s milk.
Cholesterol esterase
Another enzyme present in the pancreatic juice is cholesterol esterase (EC 3.1.1.13). The enzyme, synthesized and secreted in an active form by the exocrine pancreas, is a lipase with broad specificity, being active on:
- cholesterol esters, with cholesterol and free fatty acids as reaction products;
- triglycerides, in which it hydrolyzes all the three ester bonds, and for this reason, it is also called non-specific esterase (it hydrolyzes 10–15% of the dietary triglycerides);
- monoacylglycerols;
- phospholipids;
- esters of vitamins A and D.
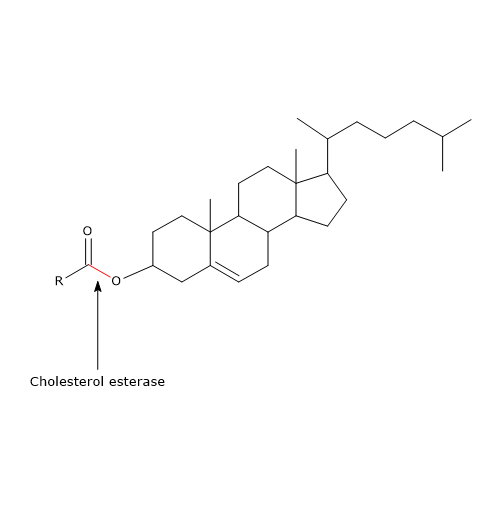
Like phospholipase A2, it is primarily active on cholesterol esters incorporated into bile salt micelles. Unlike pancreatic lipase, its activity is stimulated by bile salts, mainly trihydroxy salts, such as sodium taurocholate and glycocholate. These salts induce a protein conformational change that activates the enzyme. Moreover, trihydroxy salts promote its self-association into polymeric aggregates, which protect it from the action of proteases in the intestinal lumen.
Phospholipases
The digestion of phospholipids is carried out by phospholipases, primarily phospholipase A2 (EC 3.1.1.4). The enzyme is present in the pancreatic juice in the form of a zymogen, called prophospholipases A2, and is activated by trypsin, enzyme involved in protein digestion.
Phospholipase A2 catalyzes specifically the cleavage of the fatty acid at the sn-2 position of the phospholipids, whereas it has a broad specificity with respect to both the length of the carbon chain of the target fatty acid and the polar head groups of the phospholipids.
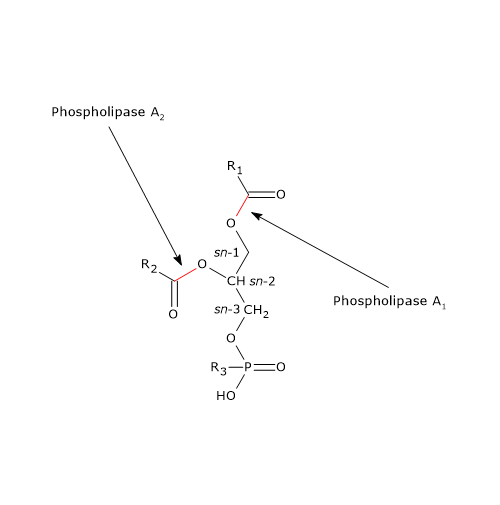
Most of the phospholipids in the intestinal lumen are of biliary origin, and only a small fraction derives from diet.
Phospholipids in bile form micelles with cholesterol and bile salts. In the intestinal lumen, they are distributed between lipid droplets and these micelles, with a preference for the latter. Within the micelles, phospholipids, primarily phosphatidylcholine, serve as a substrate. Specifically, phosphatidylcholine undergoes a reaction producing a free fatty acid and lysophosphatidylcholine, a lysophospholipid.
Pancreatic juice contains phospholipase A1 as well, which removes the fatty acid at the sn-1 position of the phospholipid.
In the intestinal mucosa, there seems to be a third, modest, phospholipase activity, thanks to an intrinsic membrane enzyme. This enzyme is called phospholipase B or retinyl ester hydrolase, being active also on vitamin A esters.
The digestion of phospholipids can end with the formation of a free fatty acid and a lysophospholipid or can be complete.
References
- Akoh C.C., Min D.B. Food lipids: chemistry, nutrition, and biotechnology. 4th Edition. Boca Raton: CRC Press, 2017. doi:10.1201/9781315151854
- Bender D.A. Introduction to nutrition and metabolism. 3rd Edition. Taylor & Francis, 2004.
- Berdanier C.D., Dwyer J., Feldman E.B. Handbook of nutrition and food. 2th Edition. CRC Press. Taylor & Francis Group, 2007
- Chow Ching K. Fatty acids in foods and their health implications. 3rd Edition. CRC Press, 2008.
- Guyton A.C., Hall J.E. Textbook of medical physiology. 14th Edition. Philadelphia: Elsevier, 2021.
- Hamosh M. Lingual and gastric lipases. Nutrition 1990;6(6):421-8.
- Iqbal J., Hussain M.M. Intestinal lipid absorption. Am J Physiol Endocrinol Metab 2009;296:E1183-1194. doi:10.1152/ajpendo.90899.2008
- Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 8th Edition. W.H. Freeman and Company, 2021.
- Rosenthal M.D., Glew R.H. Medical biochemistry – Human metabolism in health and disease. John Wiley J. & Sons, Inc., Publication, 2009.
- Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 4th Edition. St. Louis: Elsevier, 2018.
Domande Frequenti
What is the role of colipase in human lipid digestion?
Colipase is secreted as inactive procolipase and activated by trypsin. It binds to pancreatic lipase in a 1:1 molar ratio, allowing the enzyme to anchor securely to the lipid-water interface of fat droplets, preventing it from being inhibited or washed away by negatively charged bile salts.
Why is lingual lipase crucial for the digestion of newborn infants?
In newborns, pancreatic lipase is not yet fully functional. Lingual lipase compensates by effectively breaking down breast milk triglycerides, which are rich in short and medium-chain fatty acids. Unlike pancreatic lipase, it can directly penetrate and digest milk fat globules.
How does pancreatic phospholipase A2 break down phospholipids?
Secreted as a proenzyme and activated by trypsin in the intestinal lumen, phospholipase A2 specifically cleaves the fatty acid at the sn-2 position of the phospholipid. Acting inside mixed micelles, it breaks down phosphatidylcholine into a free fatty acid and lysophosphatidylcholine.