Alpha-amylase (α-amylase; EC 3.2.1.1) is an enzyme that catalyzes the hydrolysis of α-(1→4) glycosidic bonds present in the linear chains of amylose and amylopectin, the two major components of starch granules.[1][2]
It is widely distributed in animals, plants, fungi (including Ascomycota and Basidiomycota), bacteria (notably species of Bacillus), and Archaea. This broad distribution highlights its central role in carbohydrate metabolism.[3][4]
In humans, alpha-amylase is encoded by two genes and is synthesized mainly in the parotid glands and the exocrine pancreas.[5] In plants, the alpha-amylase gene family consists of six members.[6]
In animals, it initiates starch digestion, producing the disaccharide maltose, the oligosaccharides maltotriose and alpha-limit dextrins, and small amounts of glucose.[7]
Since starch is the primary dietary source of carbohydrates, and thus of glucose and energy, alpha-amylase activity may influence postprandial blood glucose levels.[8]
α-Amylase also has applications in various industrial sectors as well as in clinical diagnostics.[9][10]
In plants, particularly in grasses such as wheat, barley, and rice, it plays a crucial role in seed germination and grain maturation.[6][11]
Contents
- Historical background
- Human alpha-amylase genes
- Activity of alpha-amylase
- Salivary alpha-amylase
- Pancreatic alpha-amylase
- Blood sugar control
- Amylase activity in clinical biochemistry
- Wheat alpha-amylase
- References
Historical background
The first evidence of amylase activity dates back to 1811, when the Russian chemist Konstantin Sigizmundovich Kirchhoff, working with wheat, discovered an enzymatic activity capable of breaking down starch.[12]
In 1831, the German chemist Erhard Friedrich Leuchs observed starch degradation when mixed with human saliva, and named the responsible agent “ptyalin”.
In 1833, French chemists Anselme Payen and Jean-François Persoz, working in a sugar factory, isolated from barley an enzyme able to degrade starch, which they named diastase.[7]
The term alpha-amylase was introduced by Richard Kuhn in 1925, after he discovered that the hydrolysis products were in the alpha configuration.[13]
In 1894, alpha-amylase became the first enzyme to be produced industrially. Initially extracted from a fungal source, it was used to treat digestive disorders. Starting in 1917, it was also extracted from Bacillus subtilis and Bacillus mesentericus. Today, it is mainly produced using genetically modified organisms.[3][14]
Amylases in general, including α-amylase, beta-amylase (EC 3.2.1.2), and gamma-amylase (EC 3.2.1.3), are widely applied in industries such as textiles, detergents, animal feed, food processing, brewing (beer and whiskey), cosmetics, pharmaceuticals, and clinical diagnostics.[9]
Alpha-amylases can be classified according to the degree of substrate hydrolysis: as liquefying enzymes, when they cleave 30–40% of starch glycosidic bonds, or saccharifying enzymes, when they cleave 50–60% of the bonds.[3]
Human alpha-amylase genes
In many mammals, including humans, alpha-amylase is encoded by two distinct but closely related genes located on the short arm of chromosome 1:
- AMY1, which encodes the enzyme present in saliva, the mammary gland, and certain tumors, such as lung cancer;[15]
- AMY2, which encodes the pancreatic enzyme. AMY2 occurs in two forms, AMY2A and AMY2B, whose copy number varies among populations, although two copies of each gene appear to be common. Copy numbers of AMY2 may also be associated with those of AMY1.[5][16]
Activity of alpha-amylase
Alpha-amylase catalyzes the first step of starch digestion, the hydrolysis of α-(1→4) glycosidic bonds in the linear chains of amylose and amylopectin, by acting at random sites along α-glucans. As an endoglycosidase, it requires calcium ions, has an optimal pH around 7, and produces disaccharides and oligosaccharides in the alpha configuration. Specifically, it generates maltose and maltotriose from amylose, and maltose, alpha-limit dextrins, and small amounts of glucose from amylopectin.[17][18]
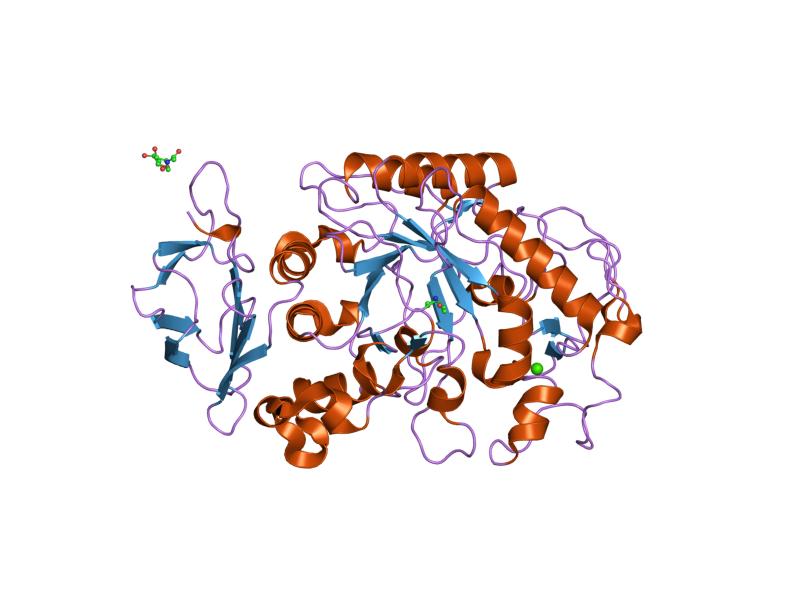
Like other enzymes involved in starch metabolism, such as pullulanase (EC 3.2.1.41), isoamylase (EC 3.2.1.68), starch-branching enzymes (EC 2.4.1.18), and starch-debranching enzymes, alpha-amylase belongs to glycoside hydrolase family 13 (GH13), a group of enzymes that act on substrates containing alpha-glycosidic bonds.[4]
Two enzymes involved in trehalose metabolism also fall within this family: trehalose synthase (EC 5.4.99.16), which catalyzes the interconversion of maltose and trehalose, and trehalose 6-phosphate hydrolase (EC 3.2.1.93), which hydrolyzes trehalose 6-phosphate to yield one molecule of glucose and one molecule of glucose 6-phosphate.[7][19]
Salivary alpha-amylase
Salivary alpha-amylase, also known as ptyalin, is synthesized mainly by the parotid glands and, to a lesser extent, by the submandibular glands.[20]
It occurs in both glycosylated and non-glycosylated forms and contributes to the initial, limited phase of starch digestion in the oral cavity.[21]
This enzyme is among the most abundant salivary proteins, accounting for 40–50% of total salivary protein content. Along with lingual lipase (EC 3.1.1.3), which hydrolyzes triglycerides and cholesterol esters during lipid digestion, salivary alpha-amylase is one of the principal enzymes in saliva.[22]
Pancreatic alpha-amylase
Pancreatic alpha-amylase is synthesized by the acinar cells of the pancreas.[23][24]
It is one of the major components of pancreatic juice and, upon entering the duodenum, hydrolyzes amylose and amylopectin, completing the first stage of amylolysis. The resulting products are subsequently cleaved by alpha-glucosidases located in the brush border of enterocytes, namely sucrase-isomaltase (EC 3.2.1.48) and maltase-glucoamylase (EC 3.2.1.20), which can also hydrolyze the α-(1→2) glycosidic bond of sucrose.[25]
Mammalian pancreatic α-amylase can bind to N-linked glycans on brush-border membrane glycoproteins.[26] This interaction enhances cooperation with sucrase-isomaltase, leading to a substantial increase in glucose production, up to 240%. However, high concentrations of alpha-amylase in the intestinal lumen have been observed to inhibit glucose uptake via SGLT1, an intestinal monosaccharide transporter involved in the absorption of monosaccharides. This inhibition has been hypothesized to represent an innate regulatory mechanism that prevents sudden spikes in glucose uptake and contributes to the regulation of postprandial blood glucose levels.[27]
Blood sugar control
Postprandial glycemic control is considered an effective strategy to prevent the development of type II diabetes. Among non-pharmacological approaches are interventions that reduce the ability of alpha-amylase to hydrolyze starch, either through food processing that induces starch retrogradation or through molecules naturally present in foods that, via different mechanisms, inhibit its activity.[8]
Food processing
Starch-rich foods are often subjected to hydrothermal treatments, such as boiling, which cause starch granule gelatinization: the granules absorb water, swell, and lose most of their crystalline structures, which are replaced by amorphous forms that are more easily hydrolyzed by alpha-amylase.[1] If gelatinized starch is subsequently cooled, recrystallization may occur, leading to the formation of retrograded starch, a type of starch resistant to α-amylase activity. This starch reaches the colon largely intact, where it is fermented by gut microbiota into short-chain fatty acids, similarly to soluble fiber.[28]
Role of polyphenols
Starch digestion by alpha-amylase is influenced by various food components, including non-starch cell wall polysaccharides, proteins, lipids, and, among phytochemicals, different classes of polyphenols.[1] These compounds can limit the action of alpha-amylase through at least four mechanisms.
- Proteins and non-starch polysaccharides can limit the loss of the ordered structure of native starch during the hydrothermal processing of foods.
- Lipids, polyphenols, proteins and non-starch polysaccharides can form ordered complexes with starch.
- Proteins, non-starch polysaccharides, and polyphenols can create physical barriers that reduce the accessibility of hydrolytic enzymes to alpha-glucan chains.
- Polyphenols and non-starch polysaccharides can directly reduce the activity of alpha-amylases and alpha-glucosidases.[8]
Among polyphenols, flavonoids, particularly proanthocyanidins (condensed tannins), appear to play a key role. For this inhibitory effect to be significant, the consumption of polyphenol-rich foods, such as black tea or green tea, should coincide with a carbohydrate-rich meal. Gluten however, may interfere with the interaction between polyphenols and starch granules, thereby reducing their inhibitory capacity.[8][29]
Amylase activity in clinical biochemistry
The measurement of serum α-amylase activity is a commonly used diagnostic laboratory test.
Increased serum amylase activity may result from:
- acute pancreatitis or an exacerbation of chronic pancreatitis;
- inflammation of the parotid glands;
- obstruction of the secretory ducts of the parotid glands or the pancreatic duct.
Current assays do not distinguish between salivary and pancreatic isoforms. However, when elevated amylase activity is accompanied by increased lipase activity, the underlying pathology is most likely of pancreatic origin.[10][30][31]
Wheat alpha-amylase
In plants, particularly in wheat, alpha-amylase plays a key role in germination and in malting, a controlled germination process that converts grain into malt, used for the production of beer and whiskey.[32]
In wheat grains, the enzyme is mainly synthesized during the early stages of germination, which are heterotrophic and rely heavily on stored resources, especially starch in the endosperm. Its synthesis occurs in the aleurone layer and initiates the mobilization of starch reserves.[11]
Alpha-amylase is also implicated in one of the major quality defects of wheat: pre-harvest sprouting.[33]
References
- ^ a b c Butterworth P.J., Bajka B.H., Edwards C.H., Warren F.J., Ellis P.R. Enzyme kinetic approach for mechanistic insight and predictions of in vivo starch digestibility and the glycaemic index of foods. Trends Food Sci Technol 2022;120:254-264. doi:10.1016/j.tifs.2021.11.015
- ^ Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 8th Edition. W.H. Freeman and Company, 2021.
- ^ a b c Tiwari S.P., Srivastava R., Singh C.S., Shukla K., Singh R.K., Singh P., Singh R., Singh N.L., and Sharma R. Amylases: an overview with special reference to alpha amylase. Journal of Global Biosciences 2015;4(SI1):1886-1901.
- ^ a b Janeček Š., Svensson B., MacGregor E.A. α-Amylase: an enzyme specificity found in various families of glycoside hydrolases. Cell Mol Life Sci 2014;71(7):1149-70. doi:10.1007/s00018-013-1388-z
- ^ a b Iafrate A.J., Feuk L., Rivera M.N., Listewnik M.L., Donahoe P.K., Qi Y., Scherer S.W, Lee C. Detection of large-scale variation in the human genome. Nat Genet 2004;36:949–951. doi:10.1038/ng1416
- ^ a b Ju L., Pan Z., Zhang H., Li Q., Liang J., Deng G., Yu M., Long H. New insights into the origin and evolution of α-amylase genes in green plants. Sci Rep 2019;9(1):4929. doi:10.1038/s41598-019-41420-w
- ^ a b c Butterworth P.J., Warren F.J. and Ellis P.R. Human α-amylase and starch digestion: an interesting marriage. Starch/Stärke 2011;63:395-405. doi:10.1002/star.201000150
- ^ a b c d Gong L., Feng D., Wang T., Ren Y., Liu Y., Wang J. Inhibitors of α-amylase and α-glucosidase: potential linkage for whole cereal foods on prevention of hyperglycemia. Food Sci Nutr 2020;8(12):6320-6337. doi:10.1002/fsn3.1987
- ^ a b de Souza P.M., de Oliveira Magalhães P. Application of microbial α-amylase in industry – A review. Braz J Microbiol 2010;41(4):850-61. doi:10.1590/S1517-83822010000400004
- ^ a b Erta G., Gersone G., Jurka A., Tretjakovs P. Salivary α-amylase as a metabolic biomarker: analytical tools, challenges, and clinical perspectives. Int J Mol Sci 2025;26(15):7365. doi:10.3390/ijms26157365
- ^ Zhang Q., Pritchard J., Mieog J., Byrne K., Colgrave M.L., Wang J.R., Ral J.F. Over-expression of a wheat late maturity alpha-amylase type 1 impact on starch properties during grain development and germination. Front Plant Sci 2022;13:811728. doi:10.3389/fpls.2022.811728
- ^ Zakowski J.J., Bruns D.E. Biochemistry of human alpha amylase isoenzymes. Crit Rev Clin Lab Sci 1985;21(4):283-322. doi:10.3109/10408368509165786
- ^ Kuhn R. Mechanism of the action of amylases. Constitution of starch. Justus Liebigs Annalen der Chemie 1925;443:1-71.
- ^ Pandey A., Nigam P., Soccol C.R., Soccol V.T., Singh D., Mohan R. Advances in microbial amylases. Biotechnol Appl Biochem 2000;31(2):135-52.
- ^ Perry G.H., Dominy N.J., Claw K.G., et al. Diet and the evolution of human amylase gene copy number variation. Nat Genet 2007;39(10):1256-60. doi:10.1038/ng2123
- ^ Carpenter D., Mitchell L.M., Armour J.A. Copy number variation of human AMY1 is a minor contributor to variation in salivary amylase expression and activity. Hum Genomics 2017;11(1):2. doi:10.1186/s40246-017-0097-3^
- ^ Benjamin S., Smitha R.B., Jisha V.N., Pradeep S., Sajith S., Sreedevi S., Priji P., Unni K.N., Sarath Josh M.K. A monograph on amylases from Bacillus spp. Adv biosci biotechnol 2013;4:No.2. doi:10.4236/abb.2013.42032
- ^ van der Maarel M.J., van der Veen B., Uitdehaag J.C., Leemhuis H., Dijkhuizen L. Properties and applications of starch-converting enzymes of the alpha-amylase family. J Biotechnol 2002;94(2):137-55. doi:10.1016/s0168-1656(01)00407-2
- ^ Agarwal N., Singh S.P. A novel trehalose synthase for the production of trehalose and trehalulose. Microbiol Spectr 2021;9(3):e0133321. doi:10.1128/Spectrum.01333-21
- ^ Schneyer L.H. Amylase content of separate salivary gland secretions of man. J Appl Physiol 1956;9(3):453-5. doi:10.1152/jappl.1956.9.3.453
- ^ Yang Z.M., Chen L.H., Zhang M., Lin J., Zhang J., Chen W.W., Yang X.R. Age differences of salivary alpha-amylase levels of basal and acute responses to citric acid stimulation between chinese children and adults. Front Physiol 2015;6:340. doi:10.3389/fphys.2015.00340
- ^ Boehlke C., Zierau O., Hannig C. Salivary amylase – The enzyme of unspecialized euryphagous animals. Arch Oral Biol 2015;60(8):1162-76. doi:10.1016/j.archoralbio.2015.05.008
- ^ Keller P.J., Allan B.J. The protein composition of human pancreatic juice. J Biol Chem 1967;242(2):281-7. doi:10.1016/S0021-9258(19)81461-8
- ^ Basnet S., Ghimire M.P., Lamichhane T.R., Adhikari R., Adhikari A. Identification of potential human pancreatic α-amylase inhibitors from natural products by molecular docking, MM/GBSA calculations, MD simulations, and ADMET analysis. PLoS One 2023;18(3):e0275765. doi:10.1371/journal.pone.0275765
- ^ Nichols B.L., Avery S., Sen P., Swallow D.M., Hahn D., Sterchi E. The maltase-glucoamylase gene: common ancestry to sucrase-isomaltase with complementary starch digestion activities. Proc Natl Acad Sci U S A. 2003 Feb 4;100(3):1432-7. doi:10.1073/pnas.0237170100
- ^ Asanuma-Date K., Hirano Y., Le N., et al. Functional regulation of sugar assimilation by N-glycan-specific interaction of pancreatic α-amylase with glycoproteins of duodenal brush border membrane. J Biol Chem 2012;287(27):23104-18. doi:10.1074/jbc.m111.314658
- ^ Date K., Satoh A., Lida K., Ogawa H. Pancreatic α-amylase controls glucose assimilation by duodenal retrieval through N-glycan-specific binding, endocytosis, and degradation. J Biol Chem 2015;290(28):17439-50. doi:10.1074/jbc.M114.594937
- ^ Holtug K., Clausen M.R., Hove H., Christiansen J., Mortensen P.B. The colon in carbohydrate malabsorption: short-chain fatty acids, pH, and osmotic diarrhoea. Scand J Gastroenterol 1992;27(7):545-52. doi:10.3109/00365529209000118
- ^ Forester S.C., Gu Y., Lambert J.D. Inhibition of starch digestion by the green tea polyphenol, (-)-epigallocatechin-3-gallate. Mol Nutr Food Res 2012;56(11):1647-54. doi:10.1002/mnfr.201200206
- ^ Ismail O.Z., Bhayana V. Lipase or amylase for the diagnosis of acute pancreatitis? Clin Biochem 2017;50(18):1275-1280. doi:10.1016/j.clinbiochem.2017.07.003
- ^ Ekka N.M., Kujur A.D., Guria R., Mundu M., Mishra B., Sekhar S., Kumar A., Prakash J., Birua H. Serum lipase amylase ratio as an indicator to differentiate alcoholic from non-alcoholic acute pancreatitis: a systematic review and meta-analysis. Cureus 2023;15(2):e35618. doi:10.7759/cureus.35618
- ^ Fincher G.B. Molecular and cellular biology associated with endosperm mobilization in germinating cereal grains. Annu Rev Plant Physiol Plant Mol Biol 1989;40:305-46. doi:10.1146/annurev.pp.40.060189.001513
- ^ Gubler F., Millar A.A., Jacobsen J.V. Dormancy release, ABA and pre-harvest sprouting. Curr Opin Plant Biol 2005;8(2):183-7. doi:10.1016/j.pbi.2005.01.011