Gli acidi grassi a catena corta (SCFA) sono acidi grassi saturi con catena lineare o ramificata composta da 2–5 atomi di carbonio, e sono gli acidi acetico, propionico, butirrico, isobutirrico, valerico, isovalerico e 2-metilbutirrico.[1]
Nell’uomo sono, con i sali biliari secondari, i principali metaboliti prodotti dai batteri del microbiota intestinale nel cieco e colon, derivando quasi interamene dalla fermentazione anerobica di carboidrati non digeribili.[2] I più abbondanti sono l’acido acetico, propionico e butirrico, che costituiscono circa il 95% del totale. La quota rimanente è costituita da quelli a catena ramificata.[3]
Sono i principali anioni presenti nel colon.[4] La loro concentrazione è maggiore nel cieco e nel colon prossimale rispetto ala parte distale, dove i substrati di sintesi si esauriscono.[5][6][7] Possono ridurre il pH del lume intestinale, acidificando le feci.
Il 90–95% viene assorbito nel cieco e nel colon, mentre il restante 5–10% viene escreto, e si ritiene forniscano circa il 70% del fabbisogno energetico necessario ai colonociti.[8][9]
Gli acidi grassi a catena corta modulano sia la fisiologia che la composizione del microbiota intestinale. Inoltre, un numero crescente di ricerche suggerisce che svolgano un ruolo importante nel supportare la salute umana.[4]
In Sintesi: Punti Chiave
- Origine microbiota: gli acidi grassi a catena corta derivano quasi interamente dalla fermentazione anaerobica di fibre e amidi resistenti da parte dei batteri intestinali.
- I tre pilastri: gli acidi acetico, propionico e butirrico sono i più abbondanti e rappresentano il 90-95% degli SCFA presenti nel cieco e nel colon.
- Carburante cellulare: l’acido butirrico è di fondamentale importanza biologica, poiché copre circa il 70% del fabbisogno energetico dei colonociti.
- Regolazione e salute: modulano la fisiologia dell’ospite agendo come inibitori delle istone deacetilasi e legandosi ai recettori di membrana GPR per regolare l’asse intestino-cervello e l’omeostasi glicemica.
Indice
- Fonti
- Proprietà
- Sintesi
- Recettori di membrana
- Assorbimento
- Trasporto
- Funzioni nel colonocita
- Metabolismo epatico ed extraepatico
- Bibliografia
Fonti
Al pari degli acidi grassi a catena media e degli acidi grassi a catena lunga, gli acidi grassi catena corta sono presenti nei tessuti animali e vegetali principalmente in forma di trigliceridi, sebbene in quantità molto inferiori rispetto a quelli a catena lunga.[10]
Nell’adulto la principale fonte alimentare è costituita da latte e derivati, dove l’acido butirrico è quello presente in concentrazione maggiore. Altre fonti sono alcuni oli vegetali, come l’olio di cocco e l’olio di palmisti.
Nei neonati allattati al seno la principale fonte è il latte materno.[11]
Tuttavia, per l’uomo, e per la maggior dei mammiferi, la fonte più importante è costituita dalla fermentazione anaerobica delle fibre e dell’amido resistente, ossia di carboidrati indigeribili, ad opera dei batteri del microbiota intestinale.[12] Questa via porta alla produzione di circa 500–600 mM di SCFA al giorno. Gli acidi acetico, propionico e butirrico sono presenti in un rapporto molare rispettivamente di circa 60:20:20, sebbene la proporzione relativa di ciascuno essi dipenda dal substrato, dalla composizione del microbiota, e dal tempo di transito intestinale.[2][4]
Proprietà
Gli acidi grassi a catena corta hanno catene carboniose composte da un numero di atomi di carbonio compreso tra 2 e 5, caratteristica che influenza fortemente le loro proprietà fisiche.
L’acido acetico, l’acido butirrico, l’acido propionico e l’acido valerico sono acidi grassi a catena lineare, mentre l’acido isobutirrico, l’acido isovalerico e l’acido 2-metil butirrico sono acidi grassi a catena ramificata.[13]
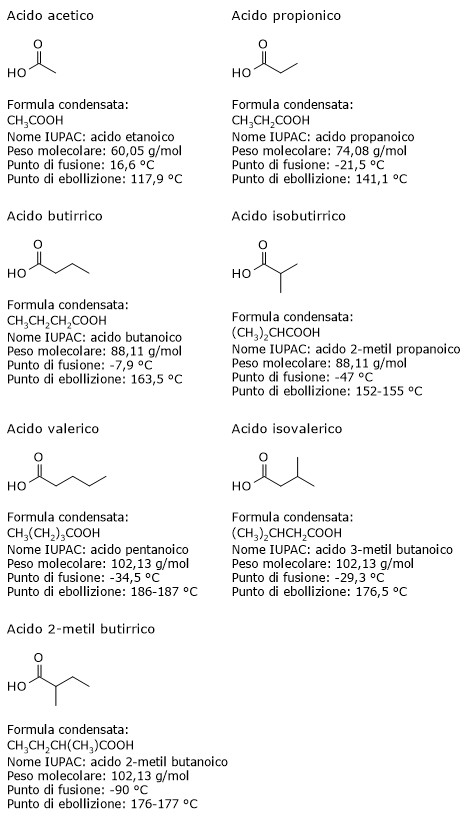
Sono molecole di piccole dimensioni, e tra i lipidi sono i più piccoli.
A temperatura ambiente sono liquidi, e sono solubili in solventi polari come l’acqua, differendo in questo dagli acidi grassi con catene carboniose più lunghe, la cui solubilità in solventi polari, considerando quelli a catena lineare, va diminuendo all’aumentare della lunghezza della catena stessa, essendo questa la parte della molecola dotata di carattere non polare, a differenza del gruppo carbossilico che è polare.[11]
Va infine notato che l’acido butirrico e l’acido isobutirrico, che hanno formula molecolare C4H8O2, sono un esempio di isomeria di catena, al pari dell’acido valerico, dell’acido isovalerico e dell’acido 2-metilbutirrico, i quali hanno formula molecolare C5H10O2.
Effetti sulla salute
Gli acidi grassi a catena corta sono ritenuti svolgere un ruolo cruciale nel mantenimento della salute dell’uomo.[7] La loro azione sembra esplicarsi attraverso effetti diretti e indiretti su processi cellulari come la proliferazione, la differenziazione e l’espressione genica, concorrendo in questo modo alla regolazione di processi quali l’omeostasi del glucosio, la funzionalità intestinale e immunitaria, e la regolazione dell’asse microbiota-intestino-cervello.[14]
Tutto ciò sembra essere confermato anche da studi che mostrano come una disbiosi intestinale appaia essere implicata in patologie metaboliche, come disturbi che coinvolgono l’omeostasi del glucosio, e comportamentali e neurologiche, come l’Alzheimer, il Parkinson, e la depressione.[15]
Sintesi
Nell’uomo, il corredo enzimatico responsabile della digestione dei carboidrati è privo degli enzimi in grado di digerire le fibre e l’amido resistente, quest’ultimo così chiamato proprio perché resiste all’azione idrolitica della alfa-amilasi. Al contrario, i batteri del microbiota intestinale codificano per un gran numero di differenti glicoside idrolasi, oltre 260, grazie alle quali sono in grado di liberare monosaccaridi dalle fibre e dall’amido resistente.[16]
Gli esosi e i desossiesosi entrano nella glicolisi, e i pentosi nella via del pentoso fosfato, a dare piruvato, che è il principale precursore per la sintesi degli acidi grassi a catena corta.[17][18][19]
La sintesi degli SCFA è influenzata da diversi fattori, alcuni dei quali sono di seguito elencati.
- Il contenuto di fibre della dieta. Ad esempio, una dieta ricca di fibre, come la dieta mediterranea, può influenzarne la sintesi.[2]
- La composizione del microbiota intestinale.[20]
- Il pH del lume intestinale, in quanto per valori di circa 5,5 dominano i batteri che producono acido butirrico, mentre per valori di circa 6,5 dominano i batteri che producono acido acetico e acido propionico.[21]
- La velocità di transito intestinale.[22]
- La quantità di ossigeno nel lume intestinale.[6]
L’acido acetico e l’acido propionico sono prodotti principalmente da specie appartenenti al phylum Bacteroidetes, mentre l’acido butirrico, per la cui sintesi è particolarmente importante l’amido resistente, da specie batteriche appartenenti al phylum Firmicutes.[23][24]
Sintesi dell’acido acetico
L’acido acetico, il più abbondante tra gli SCFA presenti nel colon, può essere sintetizzato attraverso la via di Wood-Ljungdahl nella direzione riduttiva, a mezzo della riduzione della CO2 ad acetato, o dall’acetil-CoA, la via metabolica più importante essendo responsabile della produzione di circa due terzi dell’acido grasso presente nel lume intestinale.[25]
Sintesi dell’acido propionico
L’acido propionico può essere sintetizzato attraverso tre differenti vie metaboliche: le vie dell’acrilato e del succinato, che utilizzano come precursore l’acido lattico prodotto da altri batteri, e la via del propandiolo, nella quale i precursori sono desossiesosi.[6]
Nella via dell’acrilato, l’acido lattico viene convertito in propionil-Coa via lattoil-CoA. Nel passaggio finale il propionil-Coa è idrolizzato a dare acido propionico.[26]
Nella via del succinato, il lattato viene ridotto a piruvato, che a sua volta viene carbossilato a ossalacetato, che è convertito in propionil-CoA attraverso una via metabolica che ha come intermedi il malato, il fumarato, il succinato e il metilmalonil-CoA. Infine il propionil-CoA viene idrolizzato a coenzima A e acido propionico. La via del succinato è considerata la via dominante per la sintesi dell’acido propionico nell’intestino.[27]
Nella via del propandiolo, alcuni desossiesosi, come il fucosio e il ramnosio, sono convertiti, via 1,2-propandiolo, in propionil-Coa e quindi in acido propionico.[28]
Sintesi dell’acido butirrico
La sintesi dell’acido butirrico può seguire due vie.[4][11]
Nella maggior parte dei batteri produttori di acido butirrico, l’acido grasso a catena corta è prodotto attraverso una via metabolica che ha inizio con la condensazione di due molecole di acetil-CoA a dare acetoacetil-CoA. Di seguito l’acetoacetil-CoA viene convertito in β-idrossibutirril-CoA, crotonil-CoA, e butirril-CoA. Nell’ultima tappa si ha la liberazione di acido butirrico dal butirril-CoA.[29]
In un numero ridotto di specie batteriche il butirril-CoA viene convertito in butirrilfosfato, da cui viene infine liberato acido butirrico.[6]
Sintesi dagli aminoacidi
L’acido acetico, l’acido propionico e l’acido butirrico possono essere prodotti anche a partire da aminoacidi derivanti dalla degradazione di peptidi e proteine, sebbene la quantità prodotta da queste vie sia minima.[24]
Questi processi si verificano nella parte distale del colon, spesso ad opera di batteri non commensali.[27] Dal metabolismo di aminoacidi differenti sono prodotti acidi grassi a catena corta diversi; di seguito alcuni esempi.[30]
- Dall’acido glutammico sono prodotti principalmente acido acetico e acido butirrico.
- Dall’acido aspartico derivano principalmente acido acetico e acido propionico.
- Dagli aminoacidi basici lisina, arginina e istidina sono prodotti acido acetico e acido butirrico.
- Dalla cisteina derivano gli acidi acetico, propionico e butirrico.
- Dal metabolismo della metionina derivano principalmente l’acido propionico e l’acido butirrico.
- Dagli aminoacidi ramificati leucina, isoleucina e valina si originano acidi grassi a catena corta ramificati.
Il pH del lume intestinale influenza il metabolismo delle proteine da parte del microbiota intestinale, a partire dalla loro degradazione negli aminoacidi costituenti, più probabile a pH neutro o debolmente alcalino.[31]
Si noti che dal metabolismo intestinale degli aminoacidi sono prodotti anche composti potenzialmente tossici, quali ammoniaca, solfiti e fenoli.[30]
Sintesi endogena
L’uomo, e più in generale i mammiferi, hanno il corredo enzimatico per la sintesi endogena di acidi grassi a catena corta. La sintesi, prevalentemente epatica, avviene a mezzo di cicli di reazioni di β-ossidazione che portano alla formazione di acil-CoA con catena carboniosa via via più corta dell’acido grasso di partenza. L’acido grasso legato al coenzima A è quindi rilasciato a seguito dell’idrolisi operata da una acil-CoA tioesterasi (EC 3.1.2.20).[11][32]
Recettori di membrana
Gli acidi grassi a catena corta sono in grado di legarsi a specifici recettori presenti sulla membrana plasmatica, i recettori accoppiati alle proteine G, tra cui GPR41, GPR43 e GPR109A.[33]
Gli effetti del legame ai recettori dipendono dal tipo di cellula. Ad esempio, il legame ai recettori presenti sulle cellule L dell’intestino è associato al rilascio di GLP-1, o glucagon-like peptide-1, e del peptide YY, ormoni che hanno effetto sull’appetito e sull’assunzione di cibo.[4] Il legame alle cellule enterocromaffini induce il rilascio di serotonina, il che potrebbe avere effetto sulla motilità intestinale.[34] Infine il legame ai recettori presenti sulle cellule beta del pancreas aumenta il rilascio di insulina.[35]
I diversi acidi grassi a catena corta hanno capacità differenti di attivare i recettori: GPR43 è più probabile sia attivato dall’acido acetico e dall’acido propionico, GPR41 dall’acido propionico e dall’acido butirrico, mentre GPR109A dall’acido butirrico.[36]
Assorbimento
Circa il 90% degli acidi grassi a catena corta presenti nel lume intestinale è assorbito dai colonociti.[37] Il passaggio attraverso la membrana plasmatica può avvenire per diffusione passiva o a mezzo di un trasporto attivo mediato da due tipi di trasportatori di membrana: i trasportatori per i monocarbossilati H+-dipendenti e quelli Na+-dipendenti, indicati rispettivamente come MCT e SMCT.[38]
Il trasporto passivo interessa le forme protonate degli SCFA, quindi è influenzato dal pH presente nel colon. Una debole acidificazione del lume intestinale, che può essere conseguente all’attività metabolica dei microrganismi presenti, aumenta la prevalenza della forma protonata e quindi del trasporto passivo.[11]
Funzioni nel colonocita
Nei colonociti, gli acidi grassi a catena corta possono essere utilizzati a scopi energetici e regolatori.[33]
Quando utilizzati a fini energetici, l’acido acetico e l’acido butirrico sono convertiti in acetil-CoA, e l’acido propionico in propionil-CoA. Attraverso la produzione di ATP contribuiscono al mantenimento dell’omeostasi cellulare, ma anche, ad esempio, al mantenimento dell’integrità delle giunzioni strette presenti agli apici cellulari, e quindi dell’integrità della barriera intestinale.[39] Dei tre principali acidi grassi a catena corta prodotti nell’intestino, l’acido butirrico è la principale fonte di energia per i colonociti, mentre gli acidi acetico e propionico sono scarsamente metabolizzati e per la maggior parte drenati dalla vena porta.[27][40]
A fini regolatori gli SCFA sono ad esempio in grado di inibire l’attività delle istone deacetilasi (EC 3.5.1.98), enzimi che catalizza la rimozione dei gruppi acetilici da residui di lisina delle proteine istoniche, gruppi acetilici precedentemente inseriti dalla istone acetiltransferasi (EC 2.3.1.48).[41] I gruppi R delle lisine deacetilate portano cariche positive, il che fa si che le proteine istoniche avvolgano più strettamente il DNA, che porta cariche negative. Ciò rende il nucleosoma più compatto, e di conseguenza più difficile eseguire la trascrizione e l’espressione genica. I diversi acidi grassi a catena corta hanno differenti capacità di inibire l’attività delle istone deacetilasi:
- fino all’80% per l’acido butirrico;
- fino al 60% per l’acido propionico;
- l’acido acetico ha la minor capacità inibitoria.[7]
Questa modalità d’azione sulle istone deacetilasi è stata osservata non solo nell’intestino e nel tessuto immunitario associato, ma anche nel sistema nervoso centrale e periferico.[42]
Trasporto
Gli acidi grassi a catena corta che non sono stati utilizzati nel colonocita lasciano la cellula per diffusione passiva e trasporto attivo, a livello della membrana basolaterale, per entrare nella circolazione portale, dove la concentrazione maggiore è raggiunta dall’acido acetico, circa 260 mM/L, mentre gli acidi propionico e butirrico raggiungono concentrazione di circa 30 mM/L.[4]
Nel retto, una piccola quantità di questi piccoli lipidi può passare direttamente nella circolazione sistemica, bypassando il fegato, a mezzo della vena iliaca interna.[40]
A differenza degli acidi grassi a catena lunga, quelli a catena corta e media viaggiano in circolo in forma libera, come acidi grassi non esterificati, e, legati all’albumina, raggiungono il fegato. Il successivo uptake e trasporto citosolico non richiede l’intervento delle proteine leganti gli acidi grassi, di traslocasi per gli acidi grassi nella membrana plasmatica, o di proteine leganti gli acidi grassi nel citosol. Pertanto, muovendosi liberamente, la loro ossidazione potrebbe essere assai più veloce rispetto a quella degli acidi grassi a catena lunga e ai membri più lunghi degli acidi grassi a catena media, ossia quelli con catene carboniose con più di 8 atomi.[11]
Metabolismo epatico ed extraepatico
Il fegato è un sito importante per il metabolismo degli acidi grassi a catena corta.[37]
L’organo è in grado di assorbire circa il 40% dell’acido acetico e l’80% del dell’acido propionico presenti nella vena porta.[43] L’acido propionico è per la maggior parte metabolizzato nel fegato, dove può essere utilizzato anche come substrato per la gluconeogenesi.[44]
Una piccola quantità degli SCFA prodotti nell’intestino, circa il 36% per l’acido acetico, il 9% per l’acido propionico e solo il 2% per l’acido butirrico, raggiunge, attraverso la circolazione sistemica, i tessuti periferici. Nel muscolo, l’acido acetico può essere utilizzato per la sintesi dei lipidi o essere ossidato per la produzione di energia. Inoltre, si ritiene che le concentrazioni presenti nella circolazione sistemica, seppure piccole, siano in grado di influenzare il metabolismo e la fisiologia delle cellule dei tessuti periferici.[4][12]
Bibliografia
- ^ Cummings J.H. Short chain fatty acids in the human colon. Gut 1981;22(9):763-79. doi:10.1136/gut.22.9.763
- ^ a b c Silva Y.P., Bernardi A., Frozza R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front Endocrinol (Lausanne) 2020;11:25. doi:10.3389/fendo.2020.00025
- ^ Chen X.F., Chen X., Tang X. Short-chain fatty acid, acylation and cardiovascular diseases. Clin Sci (Lond) 2020;134(6):657-676. doi:10.1042/CS20200128
- ^ a b c d e f g Portincasa P., Bonfrate L., Vacca M., De Angelis M., Farella I., Lanza E., Khalil M., Wang D.Q.-H., Sperandio M., Di Ciaula A. Gut microbiota and short chain fatty acids: implications in glucose homeostasis. Int J Mol Sci 2022;23:1105. doi:10.3390/ijms23031105
- ^ Abdul Rahim M.B.H., Chilloux J., Martinez-Gili L., Neves A.L., Myridakis A., Gooderham N., Dumas M.-E. Diet-induced metabolic changes of the human gut microbiome: importance of short-chain fatty acids, methylamines and indoles. Acta Diabetol 2019;56:493-500. doi:10.1007/s00592-019-01312-x
- ^ a b c d Deleu S., Machiels K., Raes J., Verbeke K., Vermeire S. Short chain fatty acids and its producing organisms: an overlooked therapy for IBD? EBioMedicine 2021;66:103293. doi:10.1016/j.ebiom.2021.103293
- ^ a b c Liu X.F., Shao J.H., Liao Y.T., Wang L.N., Jia Y., Dong P.J., Liu Z.Z., He D.D., Li C., Zhang X. Regulation of short-chain fatty acids in the immune system. Front Immunol 2023;14:1186892. doi:10.3389/fimmu.2023.1186892
- ^ McLoughlin R.F., Berthon B.S., Jensen M.E., Baines K.J., Wood L.G. Short-chain fatty acids, prebiotics, synbiotics, and systemic inflammation: a systematic review and meta-analysis. Am J Clin Nutr 2017;106(3):930-945. doi:10.3945/ajcn.117.156265
- ^ Brahe L.K., Astrup A., Larsen L.H. Is butyrate the link between diet, intestinal microbiota and obesity-related metabolic diseases? Obes Rev 2013;14(12):950-9. doi:10.1111/obr.12068
- ^ Marten B., Pfeuffer M., Schrezenmeir J. Medium-chain triglycerides. Int Dairy J 2006;16:11, Pages 1374-1382. doi:10.1016/j.idairyj.2006.06.015
- ^ a b c d e f Schönfeld P., Wojtczak L. Short- and medium-chain fatty acids in energy metabolism: the cellular perspective. J Lipid Res 2016;57(6):943-54. doi:10.1194/jlr.R067629
- ^ a b Cummings J.H., Pomare E.W., Branch W.J., Naylor C.P., Macfarlane G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987;28(10):1221-7. doi:10.1136/gut.28.10.1221
- ^ PubChem
- ^ Dalile B., Van Oudenhove L., Vervliet B., Verbeke K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat Rev Gastroenterol Hepatol 2019;16(8):461-478. doi:10.1038/s41575-019-0157-3
- ^ Borre Y.E., O’Keeffe G.W., Clarke G., Stanton C., Dinan T.G., Cryan J.F. Microbiota and neurodevelopmental windows: implications for brain disorders. Trends Mol Med 2014;20(9):509-18. doi:10.1016/j.molmed.2014.05.002
- ^ Usuda H., Okamoto T., Wada K. Leaky gut: effect of dietary fiber and fats on microbiome and intestinal barrier. Int J Mol Sci 2021;22(14):7613. doi:10.3390/ijms22147613
- ^ Hugenholtz F., Mullaney J. A., Kleerebezem M., Smidt H., Rosendale D. I. Modulation of the microbial fermentation in the gut by fermentable carbohydrates. Bioact Carbohydr Diet Fibre 2013;2(2)-133-142. doi:10.1016/j.bcdf.2013.09.008
- ^ Krautkramer K.A., Fan J., Bäckhed F. Gut microbial metabolites as multi-kingdom intermediates. Nat Rev Microbiol 2021;19(2):77-94. doi:10.1038/s41579-020-0438-4
- ^ Miller T.L., Wolin M.J. Pathways of acetate, propionate, and butyrate formation by the human fecal microbial flora. Appl Environ Microbiol 1996;62(5):1589-92. doi:10.1128/aem.62.5.1589-1592
- ^ Bergman E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol Rev 1990;70(2):567-90. doi: 10.1152/physrev.1990.70.2.567
- ^ Topping D.L., Clifton P.M. Short-chain fatty acids and human colonic function: roles of resistant starch and nonstarch polysaccharides. Physiol Rev 2001;81(3):1031-64. doi:10.1152/physrev.2001.81.3.1031
- ^ Macfarlane S., Macfarlane G.T. Regulation of short-chain fatty acid production. Proc Nutr Soc 2003;62:67–72. doi:10.1079/PNS2002207
- ^ Louis P., Flint H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol Lett 2009;294(1):1-8. doi: 10.1111/j.1574-6968.2009.01514.x
- ^ a b Louis P., Flint H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ Microbiol 2017;19(1):29-41. doi:10.1111/1462-2920.13589
- ^ Ragsdale S.W., Pierce E. Acetogenesis and the Wood-Ljungdahl pathway of CO2, fixation. Biochim Biophys Acta 2008;1784(12):1873-98. doi:10.1016/j.bbapap.2008.08.012
- ^ Hetzel M., Brock M., Selmer T., Pierik A.J., Golding B.T., Buckel W. Acryloyl-CoA reductase from Clostridium propionicum. An enzyme complex of propionyl-CoA dehydrogenase and electron-transferring flavoprotein. Eur J Biochem 2003;270(5):902-10. doi:10.1046/j.1432-1033.2003.03450.x
- ^ a b c Koh A., De Vadder F., Kovatcheva-Datchary P., Bäckhed F. From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell 2016;165(6):1332-1345. doi:10.1016/j.cell.2016.05.041
- ^ Scott K.P., Martin J.C., Campbell G., Mayer C.D., Flint H.J. Whole-genome transcription profiling reveals genes up-regulated by growth on fucose in the human gut bacterium Roseburia inulinivorans. J Bacteriol 2006;188(12):4340-9. doi:10.1128/JB.00137-06
- ^ Pryde S.E., Duncan S.H., Hold G.L., Stewart C.S., Flint H.J. The microbiology of butyrate formation in the human colon. FEMS Microbiol Lett 2002;217(2):133-9. doi:10.1111/j.1574-6968.2002.tb11467.x
- ^ a b Smith E.A., Macfarlane G.T. Dissimilatory amino acid metabolism in human colonic bacteria. Anaerobe 1997;3(5):327-37. doi:
- ^ Windey K., De Preter V., Verbeke K. Relevance of protein fermentation to gut health. Mol Nutr Food Res 2012;56(1):184-96. doi:10.1002/mnfr.201100542
- ^ Trefely S., Lovell C.D., Snyder N.W., Wellen K.E. Compartmentalised acyl-CoA metabolism and roles in chromatin regulation. Mol Metab 2020;38:100941. doi:10.1016/j.molmet.2020.01.005
- ^ a b Parada Venegas D., De la Fuente M.K., Landskron G., González M.J., Quera R., Dijkstra G., Harmsen H.J.M., Faber K.N., Hermoso M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front Immunol 2019;10:1486. doi:10.3389/fimmu.2019.00277
- ^ Fukumoto S., Tatewaki M., Yamada T., Fujimiya M., Mantyh C., Voss M., Eubanks S., Harris M., Pappas T.N., Takahashi T. Short-chain fatty acids stimulate colonic transit via intraluminal 5-HT release in rats. Am J Physiol Regul Integr Comp Physiol 2003;284(5):R1269-76. doi:10.1152/ajpregu.00442.2002
- ^ Puddu A., Sanguineti R., Montecucco F., Viviani G.L. Evidence for the gut microbiota short-chain fatty acids as key pathophysiological molecules improving diabetes. Mediators Inflamm 2014;2014:162021. doi:10.1155/2014/162021
- ^ Brown A.J., Goldsworthy S.M., Barnes A.A., Eilert M.M., Tcheang L., Daniels D., Muir A.I., Wigglesworth M.J., Kinghorn I., Fraser N.J., Pike N.B., Strum J.C., Steplewski K.M., Murdock P.R., Holder J.C., Marshall F.H., Szekeres P.G., Wilson S., Ignar D.M., Foord S.M., Wise A., Dowell S.J. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J Biol Chem 2003;278(13):11312-9. doi:10.1074/jbc.M211609200
- ^ a b Xiong R.G., Zhou D.D., Wu S.X., Huang S.Y., Saimaiti A., Yang Z.J., Shang A., Zhao C.N., Gan R.Y., Li H.B. Health benefits and side effects of short-chain fatty acids. Foods 2022;11(18):2863. doi:10.3390/foods11182863
- ^ Kim C.H., Park J., Kim M. Gut microbiota-derived short-chain Fatty acids, T cells, and inflammation. Immune Netw 2014;14(6):277-88. doi:10.4110/in.2014.14.6.277
- ^ Chambers E.S., Preston T., Frost G., Morrison D.J. Role of gut microbiota-generated short-chain fatty acids in metabolic and cardiovascular health. Curr Nutr Rep 2018;7(4):198-206. doi:10.1007/s13668-018-0248-8
- ^ a b van der Beek C.M., Dejong C.H.C., Troost F.J., Masclee A.A.M., Lenaerts K. Role of short-chain fatty acids in colonic inflammation, carcinogenesis, and mucosal protection and healing. Nutr Rev 2017;75(4):286-305. doi:10.1093/nutrit/nuw067
- ^ Marchix J., Goddard G., Helmrath M.A. Host-gut microbiota crosstalk in intestinal adaptation. Cell Mol Gastroenterol Hepatol 2018;6(2):149-162. doi:10.1016/j.jcmgh.2018.01.024
- ^ Stilling R.M., van de Wouw M., Clarke G., Stanton C., Dinan T.G., Cryan J.F. The neuropharmacology of butyrate: the bread and butter of the microbiota-gut-brain axis? Neurochem Int 2016;99:110-132. doi:10.1016/j.neuint.2016.06.011
- ^ Chambers E.S. Gut-derived short-chain fatty acids: a friend or foe for hepatic lipid metabolism? Nutr Bull 2019;44:154-159. doi:10.1111/nbu.12377
- ^ Leung C., Rivera L., Furness J.B., Angus P.W. The role of the gut microbiota in NAFLD. Nat Rev Gastroenterol Hepatol 2016;13(7):412-25. doi:10.1038/nrgastro.2016.85
Domande Frequenti
Cosa sono gli acidi grassi a catena corta e quali sono i principali?
Gli acidi grassi a catena corta (SCFA) sono lipidi saturi con 2-5 atomi di carbonio. I più abbondanti nel corpo umano sono l'acido acetico, il propionico e il butirrico. Rappresentano circa il 90-95% degli SCFA totali prodotti tramite la fermentazione batterica nel colon.
Come avviene la sintesi degli acidi grassi a catena corta nel colon?
Gli SCFA sono prodotti dai batteri del microbiota intestinale nel cieco e nel colon. Questo processo avviene quasi interamente attraverso la fermentazione anaerobica di carboidrati non digeribili, come le fibre alimentari e l'amido resistente introdotti con la dieta.
Quali sono le principali fonti di acidi grassi a catena corta?
La fonte principale per l'uomo è la fermentazione intestinale delle fibre. A livello alimentare, si trovano come componenti dei trigliceridi nei tessuti animali e vegetali. Negli adulti la fonte maggiore è rappresentata da latte e derivati, mentre nei neonati è il latte materno.
Qual è il ruolo dell'acido butirrico per le cellule del colon?
L'acido butirrico è la principale fonte energetica per i colonociti, le cellule del colon, coprendo circa il 70% del loro fabbisogno. Contribuisce a mantenere l'omeostasi cellulare e l'integrità delle giunzioni strette, potenziando l'efficacia della barriera intestinale.