I carotenoidi sono una classe di pigmenti liposolubili ampiamente presenti in natura. Sono composti organici di colore giallo, arancio e rosso costituiti da otto unità isopreniche e caratterizzati dalla presenza di numerosi doppi legami coniugati. La loro catena idrocarburica può subire modificazioni che ne influenzano significativamente le proprietà biologiche.[1]
Scoperti nella prima metà del 1800, i carotenoidi sono prodotti da tutti gli organismi fotosintetici, comprese piante, macroalghe, microalghe, nonché organismi non fotosintetici quali alcuni tipi di funghi, batteri e insetti come gli Afidi dei piselli, il ragnetto rosso (una specie di acaro) e alcune specie di moscerini. Nelle piante e nelle microalghe sono sintetizzati e accumulati nei plastidi.[2][3]
Con polifenoli e glucosinolati, costituiscono uno dei principali gruppi dei fitochimici.[4]
Nel corso dell’evoluzione, grazie alle loro proprietà chimiche e fisiche, si sono dimostrati estremamente versatili, essendo in grado di svolgere molte funzioni sia nelle piante che negli animali, tra le quali è molto importante l’azione antiossidante. Nell’uomo molti carotenoidi sono utilizzati come precursori della vitamina A.[5][6]
Si contano più di 750 carotenoidi differenti, più di 100 sono stati trovati nella frutta e verdura, e circa 40 sono assunti in quantità significative dall’uomo.[7][8]
Grazie al loro colore, i carotenoidi sono utilizzati come additivi alimentari.[9][10]
In Sintesi: Punti Chiave
- Natura chimica: pigmenti liposolubili (tetraterpeni) costituiti da 8 unità isopreniche con numerosi doppi legami coniugati.
- Classificazione: suddivisi in caroteni e xantofille.
- Funzioni biologiche: essenziali per la fotosintesi e la protezione cellulare tramite l’azione antiossidante.
- Salute umana: agiscono come precursori della vitamina A, supportano la funzione visiva e aiutano nella prevenzione di patologie croniche.
- Fonti e assorbimento: presenti in frutta e verdura giallo-arancio o verde scuro; nel plasma sono trasportati dalle lipoproteine e la loro biodisponibilità dipende dalla presenza di grassi nella dieta.
Indice
- Storia
- Struttura chimica
- Isomeria
- Classificazione
- Solubilità
- Colore
- Ruolo nelle piante
- Fonti alimentari
- Additivi alimentari
- Carotenoidi e salute dell’uomo
- Carotenoidi nel plasma
- Bibliografia
Storia
I carotenoidi sono stati scoperti nella prima metà del diciannovesimo secolo.
Il primo carotenoide a essere isolato fu il β-carotene nel 1831, grazie al lavoro del farmacista tedesco Heinrich Wilhelm Ferdinand Wackenroder, il quale chiamò “carotene” il pigmento giallo che riuscì a cristallizzare dalla radice della carota.[2]
Jöns Jacob Berzelius, considerato uno dei padri della chimica moderna, chiamò i pigmenti gialli estratti dalle foglie autunnali “xantofille”, dal greco xanthos (giallo), e phyllon (foglia).[11][12]
Mikhail Semyonovich Tsvet, botanico russo che nel 1906 ideò la cromatografia, riuscì a separare i pigmenti fogliari, ossia le clorofille dai caroteni e xantofille, grazie alla cromatografia su colonna, e chiamò carotenoidi i pigmenti dal giallo all’arancio.[1]
Struttura chimica
I carotenoidi sono una classe di lipidi aventi come unità strutturale di base una sequenza lineare di otto unità isopreniche. Sono quindi terpeni, precisamente tetraterpeni, per un totale di 40 atomi di carbonio.[8]
Le unità isopreniche sono unite con una relazione posizionale 1,5 (testa-coda), tranne al centro della catena idrocarburica, dove la relazione posizionale è 1,6 (coda-coda). Questa inversione rende la molecola simmetrica.[5]
I carotenoidi sono caratterizzati dalla presenza di un esteso sistema di doppi legami, di cui possono essere coniugati da 3 a 13.[13]
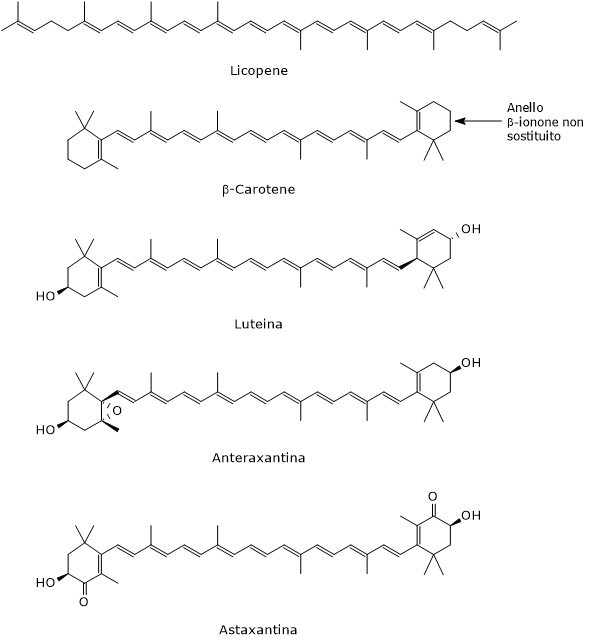
La catena idrocarburica dei carotenoidi può andare incontro a modificazioni chimiche quali idrogenazione, deidrogenazione, introduzione di atomi di ossigeno, ciclizzazione di una o entrambe le estremità a formare anelli ionone, ed esterificazione con acidi grassi.[14] Queste modificazioni permettono la formazione di molte strutture differenti, la maggior parte delle quali sono descritte dalla formula chimica C40H56On, con 0 ≤ n ≤ 6.[7]
Tradizionalmente, ai carotenoidi sono stati assegnati nomi comuni legati alla fonte biologica da cui sono stati originariamente isolati, come β-carotene (dalla carota) o astaxantina (dall’aragosta), sebbene possano anche essere denominati utilizzando la nomenclatura IUPAC.[6][15]
Isomeria
I carotenoidi presentano molti doppi legami che possono avere isomeria cis–trans, anche detta isomeria geometrica. Sebbene, in teoria, ognuno dei doppi legami possa essere in configurazione cis o trans, in natura la configurazione prevalente è la trans, essendo termodinamicamente più stabile rispetto alla configurazione cis grazie al minor ingombro sterico tra i sostituenti.[5]
La rotazione intorno ai legami semplici, o legami sigma (σ), presenti lungo la catena carboniosa permette anche la formazione di isomeri conformazionali. Quando i doppi legami sono presenti sullo stesso lato rispetto al legame sigma si parla di conformazione s-cis. Al contrario, se i doppi legami sono su lati opposti rispetto al legame σ si parla di conformazione s-trans.[16]
In natura i carotenoidi aciclici sono principalmente in conformazione s-trans, in quanto è quella a minor ingombro sterico.[17] In presenza di strutture cicliche alle estremità della catena del carotenoide, la rotazione intorno al legame sigma C6-C7 da luogo alla formazione di un isomero 6-s-cis a minor ingombro sterico e quindi energeticamente favorito.[18]
Si noti che l’ingombro sterico e le dimensioni dei carotenoidi influenzano la capacità di interagire con enzimi e di formare strutture sovramolecolari.[19]
La maggior parte dei carotenoidi presenta anche un centro di chiralità o un asse di chiralità, che fa si che esistano isomeri ottici. In questi casi, per dare un nome univoco alla molecola si ricorre al sistema RS, e non alle regole della convenzione di Fischer, utilizzata solo per carboidrati e aminoacidi.[20]
Classificazione
I carotenoidi possono essere classificati sulla base della presenza o meno di ossigeno o di strutture cicliche.[14]
Sulla base della presenza o meno di ossigeno, sono suddivisi in xantofille e caroteni.
- I caroteni, privi di ossigeno, sono formati da soli atomi di carbonio e idrogeno e hanno formula chimica C40H56. Esempi di caroteni sono l’α-carotene, il β-carotene, il δ-carotene, lo ζ-carotene, il fitoene, e il licopene.
- Le xantofille contengono anche atomi di ossigeno, hanno formula chimica C40H56On, con 1 ≤ n ≤ 6, e possono a loro volta essere suddivise in:
- idrossicarotenoidi, che contengono almeno un gruppo idrossilico, come l’α-criptoxantina, la β-criptoxantina, la zeaxantina e la luteina;
- epossicarotenoidi, che presentano almeno un gruppo epossidico, come l’anteraxantina, l’auroxantina e la luteoxantina;
- chetocarotenoidi, che presentano uno o più gruppi carbonilici, come l’astaxantina e la cantaxantina.[8]
Sulla base della presenza o meno di strutture cicliche i carotenoidi possono essere suddivisi in ciclici e aciclici.
- I carotenoidi ciclici contengono una o due strutture cicliche, e, rispetto agli aciclici, sono più corti ma con un ingombro sterico maggiore. Esempi sono l’α-carotene, il β-carotene, il γ-carotene e il δ-carotene.
- I carotenoidi aciclici sono molecole formate da una catena carboniosa lineare. Esempi sono il licopene, lo ζ-carotene, il fitoene, e il fitofluene.
Infine, esistono anche carotenoidi non comuni o specie-specifici, come la bixina, la capsantina e la capsorubina.[12]
Solubilità
I carotenoidi sono molecole idrofobiche, quindi solubili in solventi organici. Tale solubilità varia in funzione dei sostituenti presenti.[21] A causa della loro natura idrofobica, nelle cellule si ritrovano nelle membrane cellulari. Nella maggior parte dei casi, le xantofille presentano i gruppi polari alle estremità della catena, e, quando presenti nelle membrane cellulari, al fine di minimizzare l’energia del sistema, si dispongono in modo tale che i gruppi polari siano in contato con i gruppi polari del doppio strato lipidico.[22]
I carotenoidi possono avere accesso anche all’ambiente acquoso se in associazione a proteine cui si legano in modo non covalente. Nel plasma sono trasportati dalle lipoproteine.[23]
Colore
Il sistema di doppi legami coniugati, agendo come un cromoforo, è responsabile del colore dei carotenoidi.[14] Tuttavia, perché il colore sia percettibile, devono essere presenti almeno sette doppi legami coniugati, per cui molecole come il fitofluene, che ha cinque legami coniugati, o il fitoene, che ne ha tre, sono incolori.[24]
All’aumentare del numero dei doppi legami coniugati, il colore vira dal giallo, all’arancio, fino al rosso. Ad esempio, la luteina, l’α-criptoxantina e la violaxantina sono gialle, l’α-carotene, il β-carotene, e il γ-carotene sono arancioni, e il licopene è rosso.[1]
Ruolo nelle piante
Nelle piante i carotenoidi svolgono molte funzioni.
Contribuiscono alla protezione della cellula nei confronti del danno ossidativo, aumentano l’assorbimento della luce solare, sono i precursori di fitormoni, come l’acido abscissico, e intervengono nelle vie di segnalazione.[5]
Concorrono, con la clorofilla e gli antociani (una classe di flavonoidi), a determinare il colore delle foglie, dei frutti, delle verdure, dei cereali. Questa pigmentazione che ha la funzione di attirare gli animali così da promuovere l’impollinazione e la dispersione dei semi.[25][26]
Possono infine fungere anche da agenti repellenti per fitofagi e patogeni.[27]
Quenching dell’ossigeno singoletto
Negli organismi che svolgono la fotosintesi ossigenica, i carotenoidi contribuscono alla protezione dai danni dovuti all’eccessiva irradiazione solare.[28]
Quando la luce che raggiunge il complesso antenna del fotosistema II eccede la capacità di utilizzazione dei centri di reazione, l’energia in eccesso può far sì che la clorofilla resti nello stato eccitato, favorendo la formazione del suo stato di tripletto. Tale energia può essere trasferita dalla clorofilla all’ossigeno molecolare, generando ossigeno singoletto, una specie reattiva in grado di danneggiare l’apparato fotosintetico e, più in generale, acidi nucleici, proteine, acidi grassi insaturi di membrana e cofattori enzimatici.[29]
I carotenoidi possono prevenire il danno ossidativo causato dall’ossigeno singoletto in due modi.
- Possono reagire direttamente con l’ossigeno singoletto, in un processo noto come quenching. In questo caso l’energia viene trasferita al carotenoide; l’ossigeno torna allo stato fondamentale di tripletto, mentre il carotenoide passa allo stato di tripletto e dissipa l’energia in eccesso sotto forma di calore. La capacità di disattivazione aumenta con il numero di doppi legami coniugati, divenendo massima da nove o più doppi legami coniugati.
Studi in vitro indicano che i carotenoidi monociclici disattivano l’ossigeno singoletto più efficacemente rispetto alle strutture acicliche, e che la presenza di gruppi carbonilici, come nell’astaxantina, incrementa il potenziale antiossidante rispetto ai carotenoidi con gruppi idrossilici, probabilmente grazie al più esteso sistema di doppi legami coniugati. Ogni molecola di carotenoide sembra poter disattivare circa 1.000 molecole di ossigeno singoletto prima di degradarsi.[30] - I carotenoidi possono prevenire la formazione dell’ossigeno singoletto assorbendo l’energia della clorofilla eccitata, passando a loro volta allo stato di tripletto. Poiché il loro livello energetico è inferiore a quello necessario per generare ossigeno singoletto, decadono spontaneamente allo stato fondamentale.[31]
Scavenging dei radicali liberi
I carotenoidi sono in grado di agire da scavenger dei radicali liberi, quali il radicale idrossilico, i radicali perossidici, il radicale superossido e il radicale ossido nitrico. La loro azione può avvenire secondo quattro meccanismi differenti.[8][32]
- Trasferimento elettronico
I carotenoidi possono donare un elettrone al radicale, a formare il catione radicale carotenoide mentre il radicale attaccante diventa una specie stabile. Il catione radicale carotenoide può essere rigenerato nel carotenoide di partenza grazie all’azione di altri antiossidanti cellulari come la vitamina E, la vitamina C e il glutatione.[33] - Trasferimento protonico
Un protone può essere trasferito dal carotenoide al radicale, che diventa una specie neutra, mentre il carotenoide diviene un anione radicale.[34] - Reazioni di addizione
La neutralizzazione del radicale può avvenire a seguito di reazioni di addizione, come nel caso della neutralizzazione dei radicali idrossilici e perossidici.[13] - Trasferimento di atomi di idrogeno
Un atomo di idrogeno può essere trasferito dal carotenoide al radicale, con formazione di un radicale carotenoide stabilizzato per risonanza.[19]
Ruolo fotoassorbente
I carotenoidi sono presenti nei complessi antenna in qualità di pigmenti fotoassorbenti accessori, principalmente associati alle proteine antenna.[35]
Grazie al loro sistema di doppi legami coniugati, sono in grado di assorbire la luce solare nell’intervallo 400−500 nm. L’energia assorbita viene quindi convogliata ai centri di reazione.
Gli spettri di assorbimento dei singoli carotenoidi dipendono dalla lunghezza del sistema coniugato e dal tipo di gruppi funzionali presenti.[17]
Il principale carotene coinvolto nell’assorbimento della radiazione solare è il β-carotene, presente nel nucleo del fotosistema I e II di tutti gli organismi che operano una fotosintesi ossigenica, mentre la luteina, la violaxantina, la neoxantina e la zeaxantina, le principali xantofille nelle piante, sono legate ai complessi antenna.[36]
Fonti alimentari
Gli esseri umani non sono in grado di sintetizzare i carotenoidi. Li assumono con gli alimenti, accumulandoli principalmente nel fegato e nel tessuto adiposo, ma anche nei polmoni, reni, cervello e ossa, dove svolgono molte funzioni.[13]
Di tutti i carotenoidi attualmente identificati, un centinaio sono presenti negli alimenti consumati dall’uomo, mentre, se si considera il singolo alimento, in genere ne sono presenti da uno a cinque principali, accompagnati da altri presenti solo in tracce o piccole quantità.[37]
Frutta e verdura sono la principale fonte di carotenoidi nella dieta umana. Tra le verdure, quelle a foglia verde sono ricche di β-carotene, il carotenoide più abbondante nella dieta e nei tessuti umani, e di luteina, seguiti da violaxantina e neoxantina. La radice della carota è ricca di β-carotene, la zucca di α-carotene, il pomodoro di licopene, e i peperoni di capsantina e capsorubina. I carotenoidi della frutta sono molto vari, e quelli dei frutti maturi possono essere diversi da quelli dei frutti acerbi. Inoltre, la maggior parte dei carotenoidi presenti nei frutti maturi sono esterificati con acidi grassi. Tuttavia, quelli di alcuni frutti, in particolare dei frutti che a maturazione terminata rimangono verdi, come il kiwi, subiscono una limitata o assente esterificazione.[21]
Le uova di gallina, grazie al fatto che le galline ovaiole mangiano per la maggior parte il mais, possono essere una buona fonte di alcuni carotenoidi, come la luteina e zeaxantina, responsabili del colore giallo-arancio del tuorlo.[38]
Infine, molluschi e crostacei, assieme ai pesci sono le fonti sono la principale fonte dei carotenoidi prodotti dalle microalghe e raramente presenti nelle piante, come l’astaxantina, che si accumula nei gamberetti e contribuisce alla colorazione del salmone.[39]
Additivi alimentari
I carotenoidi utilizzati come additivi nell’industria alimentare, a differenza dei carotenoidi naturalmente presenti, sono relativamente instabili, essendo suscettibili all’azione dell’ossigeno e della luce solare, all’auto-ossidazione, e alla dispersione nell’alimento, che ne facilita la degradazione.[21] Inoltre, alla temperatura corporea e a quelle di conservazione, i carotenoidi aggiunti sono presenti in forma cristallina a causa del loro elevato punto di fusione. Per ovviare a questo problema, vengono utilizzati in forma di emulsione olio in acqua.[40]
Grazie al loro potere colorante, sono utilizzati come coloranti in prodotti che a causa della lavorazione o della conservazione tendono a perdere parte del loro colore naturale, o per uniformare il colore del prodotto, come nel caso delle bevande, dei succhi di frutta o degli insaccati, o per intensificarne il colore migliorandone l’aspetto.[41]
I carotenoidi, essendo precursori di composti responsabili del sapore e aroma di alcuni alimenti, possono essere utilizzati come esaltatori di sapidità.[42]
Poiché alcuni carotenoidi sono precursori della vitamina A, sono utilizzati per fortificare gli alimenti.[43]
Va infine sottolineato che, quando aggiunti agli alimenti, non sono potenti antiossidanti.[44]
Carotenoidi e salute dell’uomo
Nell’uomo i carotenoidi svolgono importanti funzioni.[45]
Il loro ruolo principale è quello di precursori della vitamina A, che da sola regola l’espressione di quasi 700 geni. Non tutti i carotenoidi lo sono: solo quelli con un anello β-ionone non sostituito. Il più importante è il β-carotene, la provitamina A per eccellenza, che possiede due anelli β-ionone non sostituiti e può quindi essere convertito in due molecole di vitamina A. L’α-carotene e la β-criptoxantina, avendo un solo anello β-ionone non sostituito, generano una sola molecola di vitamina A e hanno quindi il 50% di attività provitaminica del β-carotene.[13]
L’importanza di questa attività è evidenziata dal fatto che la carenza di vitamina A può portare a cecità notturna e xeroftalmia, al rallentamento della crescita e della rigenerazione delle mucose, all’aumento del tasso di mortalità causato dalle malattie infettive per alterazioni della risposta immunitaria, ed è la principale causa di cecità infantile nei paesi in via di sviluppo.[46]
I carotenoidi contribuiscono alla salute anche concorrendo alla prevenzione di numerose patologie croniche attraverso meccanismi diversi rispetto all’azione di provitamina A, come l’azione antiossidante e antinfiammatoria e la fotoprotezione. Di seguito alcuni esempi.
- È stato osservato che la supplementazione con luteina e zeaxantina, due xantofille che si accumulano nella macula lutea, è associata a un miglioramento della funzione visiva e a una riduzione del rischio di progressione della degenerazione maculare senile. Luteina e zeaxantina sembrano avere anche un effetto preventivo sullo sviluppo della cataratta e di retinopatie, e la luteina può avere un effetto positivo sulle funzioni cognitive.[47]
- Tra i caroteni, il consumo di alimenti ricchi di licopene è stato associato a una riduzione dell’incidenza di certi tipi di tumore, come quelli della prostata e dello stomaco, e a una riduzione del rischio cardiovascolare.[48]
Carotenoidi nel plasma
α-Carotene, β-carotene, licopene, luteina, β-criptoxantina e zeaxantina costituiscono oltre il 90% dei carotenoidi presenti nel plasma.[49]
Le concentrazioni plasmatiche dei carotenoidi dipendono da diversi fattori tra cui i metodi di cottura dei cibi, la quantità di lipidi nella dieta e variabili individuali riguardanti i processi di digestione dei lipidi e assorbimento dei lipidi medesimi, con i sali biliari a svolgere un ruolo essenziale. Anche i livelli di vitamina A nell’organismo influenzano i livelli plasmatici: un soggetto che abbia bassi livelli di vitamina A avrà probabilmente tassi di conversione dei carotenoidi precursori della vitamina A più elevati, e ciò potrebbe riflettersi in una loro minore concentrazione plasmatica.[50][51]
Bibliografia
- ^ a b c Maoka T. Carotenoids as natural functional pigments. J Nat Med 2020;74(1):1-16. doi:10.1007/s11418-019-01364-x
- ^ a b Sourkes T.L. The discovery and early history of carotene. Bull Hist Chem 2009;34(1):32-37.
- ^ Aizpuru A., González-Sánchez A. Traditional and new trend strategies to enhance pigment contents in microalgae. World J Microbiol Biotechnol 2024;40(9):272. doi:10.1007/s11274-024-04070-3
- ^ de la Rosa L.A., Alvarez-Parrilla E., Gonzàlez-Aguilar G.A. Fruit and vegetable phytochemicals: chemistry, nutritional value, and stability. 1st Edition. Wiley J. & Sons, 2010.
- ^ a b c d Swapnil P., Meena M., Kumar Singh S., Praveen Dhuldhaj U., Harish, Marwal A. Vital roles of carotenoids in plants and humans to deteriorate stress with its structure, biosynthesis, metabolic engineering and functional aspects. Curr Plant Biol 2021;26: 100203. doi:10.1016/j.cpb.2021.100203
- ^ a b Crupi P., Faienza M.F., Naeem M.Y., Corbo F., Clodoveo M.L., Muraglia M. Overview of the potential beneficial effects of carotenoids on consumer health and well-being. Antioxidants (Basel) 2023;12(5):1069. doi:10.3390/antiox12051069
- ^ a b Desmarchelier C., Borel P. Overview of carotenoid bioavailability determinants: from dietary factors to host genetic variations. Trends Food Sci Technol 2017;69(Part B):270-280. doi:10.1016/j.tifs.2017.03.002
- ^ a b c d Rodriguez-Amaya D.B. Food carotenoids: chemistry, biology, and technology. Hoboken (NJ): Wiley-Blackwell; 2015. doi:10.1002/9781118864364
- ^ Gebhardt B., Sperl R., Carle R., Müller-Maatsch J. Assessing the sustainability of natural and artificial food colorants. J Clean Prod 2020;260;120884. doi:10.1016/j.jclepro.2020.120884
- ^ EFSA Food colours. Last reviewed date: 14 November 2025.
- ^ Olson J.A., Krinsky N.I. Introduction: the colorful, fascinating world of the carotenoids: important physiologic modulators. FASEB J 1995;9(15):1547-50. doi:10.1096/fasebj.9.15.8529833
- ^ a b Britton G., Liaaen-Jensen S., Pfander H. Carotenoids: Handbook. Basel: Birkhäuser; 2004. doi:10.1007/978-3-0348-7836-4
- ^ a b c d Saini R.K., Prasad P., Lokesh V., Shang X., Shin J., Keum Y.S., Lee J.H. Carotenoids: dietary sources, extraction, encapsulation, bioavailability, and health benefits – A review of recent advancements. Antioxidants 2022;11(4):795. doi:10.3390/antiox11040795
- ^ a b c Zia-Ul-Haq M., Dewanjee S., Riaz M. Carotenoids: structure and function in the human body. Springer; 2021. doi:10.1007/978-3-030-46459-2_1
- ^ Yabuzaki J. Carotenoids database: structures, chemical fingerprints and distribution among organisms. Database (Oxford) 2017;2017(1):bax004. doi:10.1093/database/bax004
- ^ Solomons T.W.G., Fryhle C.B., Snyder S.A. Solomons’ organic chemistry. 12th Edition. John Wiley & Sons Incorporated, 2017.
- ^ a b Telegina T.A., Vechtomova Y.L., Aybush A.V., Buglak A.A., Kritsky M.S. Isomerization of carotenoids in photosynthesis and metabolic adaptation. Biophys Rev 2023;15(5):887-906. doi:10.1007/s12551-023-01156-4
- ^ Britton G. Structure and properties of carotenoids in relation to function. FASEB J 1995;9(15):1551-8. doi:10.1096/fasebj.9.15.8529834
- ^ a b Polyakov N.E., Focsan A.L., Gao Y., Kispert L.D. The endless world of carotenoids-Structural, chemical and biological aspects of some rare carotenoids. Int J Mol Sci 2023;24(12):9885. doi:10.3390/ijms24129885
- ^ Prelog V., Helmchen G. Basic principles of the CIP‐system and proposals for a revision. Angew Chem 1982:21(8);567-583. doi:10.1002/anie.198205671
- ^ a b c González-Peña M.A., Ortega-Regules A.E., Anaya de Parrodi C., Lozada-Ramírez J.D. Chemistry, occurrence, properties, applications, and encapsulation of carotenoids-A review. Plants (Basel) 2023;12(2):313. doi:10.3390/plants12020313
- ^ Gruszecki W.I., Strzałka K. Carotenoids as modulators of lipid membrane physical properties. Biochim Biophys Acta 2005;1740(2):108-15. doi:10.1016/j.bbadis.2004.11.015
- ^ Tyssandier V., Choubert G., Grolier P., Borel P. Carotenoids, mostly the xanthophylls, exchange between plasma lipoproteins. Int J Vitam Nutr Res 2002;72(5):300-8. doi:10.1024/0300-9831.72.5.300
- ^ Rodriguez-Amaya D.B., Esquivel P., Meléndez-Martínez A.J. Comprehensive update on carotenoid colorants from plants and microalgae: challenges and advances from research laboratories to industry. Foods 2023;12(22):4080. doi:10.3390/foods12224080
- ^ Archetti, M., Döring T.F., Hagen S.B., Hughes N.M., Leather S.R., Lee D.W., Lev-Yadun S., Manetas Y., Ougham H.J. Unravelling the evolution of autumn colours: an interdisciplinary approach. Trends Ecol Evol 2009;24(3):166-73. doi:10.1016/j.tree.2008.10.006
- ^ Cazzonelli C.I. Carotenoids in nature: insights from plants and beyond. Funct Plant Biol 2011;38(11):833-847.
- ^ Gómez-Sagasti M.T., López-Pozo M., Artetxe U., Becerril J.M., Hernández A., García-Plazaola J.I., Esteban R. Carotenoids and their derivatives: a “swiss army knife-like” multifunctional tool for fine-tuning plant-environment interactions. Environ Exp Bot 2023;207:105229. doi:10.1016/j.envexpbot.2023.105229
- ^ Havaux M. Carotenoids as membrane stabilizers in chloroplasts. Trends Plant Sci 1998;3(4):147-151. doi:10.1016/S1360-1385(98)01200-X
- ^ Triantaphylidès C., Havaux M. Singlet oxygen in plants: production, detoxification and signaling. Trends Plant Sci 2009;14(4):219-28. doi:10.1016/j.tplants.2009.01.008
- ^ Ramel F., Birtic S., Cuiné S., Triantaphylidès C., Ravanat J.L., Havaux M. Chemical quenching of singlet oxygen by carotenoids in plants. Plant Physiol 2012;158(3):1267-78. doi:10.1104/pp.111.182394
- ^ Pérez-Gálvez A., Viera I., Roca M. Carotenoids and chlorophylls as antioxidants. Antioxidants (Basel) 2020;9(6):505. doi:10.3390/antiox9060505
- ^ Krinsky N.I. Antioxidant functions of carotenoids. Free Radic Biol Med 1989;7(6):617-35. doi:10.1016/0891-5849(89)90143-3
- ^ Ribeiro D., Freitas M., Silva A.M.S., Carvalho F., Fernandes E. Antioxidant and pro-oxidant activities of carotenoids and their oxidation products. Food Chem Toxicol 2018;120:681-699. doi:10.1016/j.fct.2018.07.060
- ^ Focsan A.L., Kispert L.D. Radicals formed from proton loss of carotenoid radical cations: a special form of carotenoid neutral radical occurring in photoprotection. J Photochem Photobiol B 2017;166:148-157. doi:10.1016/j.jphotobiol.2016.11.015
- ^ Cogdell R.J., Gillbro T., Andersson P.O., Liu R.S.H., Asato A.E. Carotenoids as accessory light-harvesting pigments. Pure Appl Chem 1994:66(5):1041-1046. doi:10.1351/pac199466051041
- ^ Xu P., Chukhutsina V.U., Nawrocki W.J., Schansker G., Bielczynski L.W., Lu Y., Karcher D., Bock R., Croce R. Photosynthesis without β-carotene. eLife 2020;9:e58984. doi:10.7554/eLife.58984
- ^ Meléndez-Martínez A.J., Mandić A.I., Bantis F., et al. A comprehensive review on carotenoids in foods and feeds: status quo, applications, patents, and research needs. Crit Rev Food Sci Nutr 2022;62(8):1999-2049. doi:10.1080/10408398.2020.1867959
- ^ Ko E.Y., Saini R.K., Keum Y.S., An B.K. Age of laying hens significantly influences the content of nutritionally vital lipophilic compounds in eggs. Foods 2020;10(1):22. doi:10.3390/foods10010022
- ^ Galasso C., Corinaldesi C., Sansone C. Carotenoids from marine organisms: biological functions and industrial applications. Antioxidants (Basel) 2017;6(4):96. doi:10.3390/antiox6040096
- ^ Kirkhus B., Afseth N.K., Borge G.I.A., Grimsby S., Steppeler C., Krona A., Langton M. Increased release of carotenoids and delayed in vitro lipid digestion of high pressure homogenized tomato and pepper emulsions. Food Chem 2019;285:282-289. doi:10.1016/j.foodchem.2019.01.124
- ^ Mezzomo N., Ferreira S.R.S. Carotenoids functionality, sources, and processing by supercritical technology: a review. J Chem 2016;1:Article ID 3164312. doi:10.1155/2016/3164312
- ^ Meléndez-Martínez A.J., Esquivel P., Rodriguez-Amaya D.B. Comprehensive review on carotenoid composition: transformations during processing and storage of foods. Food Res Int 2023;169:112773. doi:10.1016/j.foodres.2023.112773
- ^ Hombali A.S., Solon J.A., Venkatesh B.T., Nair N.S., Peña-Rosas J.P. Fortification of staple foods with vitamin A for vitamin A deficiency. Cochrane Database Syst Rev 2019;5(5):CD010068. doi:10.1002/14651858.CD010068.pub2
- ^ Terao J. Revisiting carotenoids as dietary antioxidants for human health and disease prevention. Food Funct 2023;14:7799-7824. doi:10.1039/d3fo02330c
- ^ Eggersdorfer M., Wyss A. Carotenoids in human nutrition and health. Arch Biochem Biophys 2018;652:18-26. doi:10.1016/j.abb.2018.06.001
- ^ Hodge C., Taylor C. Vitamin A deficiency. [Updated 2023 Jan 2]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK567744/
- ^ Mrowicka M., Mrowicki J., Kucharska E., Majsterek I. Lutein and zeaxanthin and their roles in age-related macular degeneration-neurodegenerative disease. Nutrients 2022;14(4):827. doi:10.3390/nu14040827
- ^ Khan U.M., Sevindik M., Zarrabi A., et al. Lycopene: food sources, biological activities, and human health benefits. Oxid Med Cell Longev 2021;2021:2713511. doi:10.1155/2021/2713511
- ^ Deming D.M., Erdman Jr J.W. Mammalian carotenoid absorption and metabolism. Pure Appl Chem 1999;71(12):2213-2223.
- ^ Burrows T.L., Williams R., Rollo M., Wood L., Garg M.L., Jensen M., Clare Collins C.E. Plasma carotenoid levels as biomarkers of dietary carotenoid consumption: a systematic review of the validation studies. J Nutr Intermed Metab 2015;2(1-2):15-64. doi:10.1016/j.jnim.2015.05.001
- ^ Böhm V., Lietz G., Olmedilla-Alonso B., et al. From carotenoid intake to carotenoid blood and tissue concentrations – implications for dietary intake recommendations. Nutr Rev 2021;79(5):544-573. doi:10.1093/nutrit/nuaa008