Lignans are a subgroup of non-flavonoid polyphenols.
They are widely distributed in the plant kingdom, being present in more than 55 plant families, where they act as antioxidants and defense molecules against pathogenic fungi and bacteria.
In humans, epidemiological and physiological studies have shown that they can exert positive effects in the prevention of lifestyle-related diseases, such as type II diabetes and cancer. For example, an increased dietary intake of these polyphenols correlates with a reduction in the occurrence of certain types of estrogen-related tumors, such as breast cancer in postmenopausal women.
In addition, some lignans have also aroused pharmacological interest. Examples are:
- podophyllotoxin, obtained from plants of the genus Podophyllum (Berberidaceae family); it is a mitotic toxin whose derivatives have been used as chemotherapeutic agents;
- arctigenin and tracheologin, obtained from tropical climbing plants; they have antiviral properties and have been tested in the search for a drug to treat AIDS .
Contents
Chemical structure
Their basic chemical structure consists of two phenylpropane units linked by a C-C bond between the central atoms of the respective side chains (position 8 or β), also called β-β’ bond. 3-3′, 8-O-4′, or 8-3′ bonds are observed less frequently; in these cases the dimers are called neolignans. Hence, their chemical structure is referred to as (C6-C3)2, and they are included in the phenylpropanoid group, as well as their precursors: the hydroxycinnamic acids.
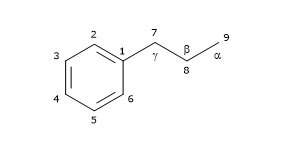
Based on their carbon skeleton, cyclization pattern, and the way in which oxygen is incorporated in the molecule skeleton, they can be divided into 8 subgroups: furans, furofurans, dibenzylbutanes, dibenzylbutyrolactones, dibenzocyclooctadienes, dibenzylbutyrolactols, aryltetralins and arylnaphthalenes. Furthermore, there is considerable variability regarding the oxidation level of both the propyl side chains and the aromatic rings.
They are not present in the free form in nature, but linked to other molecules, mainly as glycosylated derivatives.
Among the most common lignans, secoisolariciresinol (the most abundant one), lariciresinol, pinoresinol, matairesinol and 7-hydroxymatairesinol are found.
Note: They occur not only as dimers but also as more complex oligomers, such as dilignans and sesquilignans.
Biosynthesis
In this section, we will examine the biosynthesis of some of the most common lignans.
The pathway starts from 3 of the 4 most common dietary hydroxycinnamic acids: p-coumaric acid, sinapic acid, and ferulic acid (caffeic acid is not a precursor of this subgroup of polyphenols). Therefore, they arise from the shikimic acid pathway, via phenylalanine.
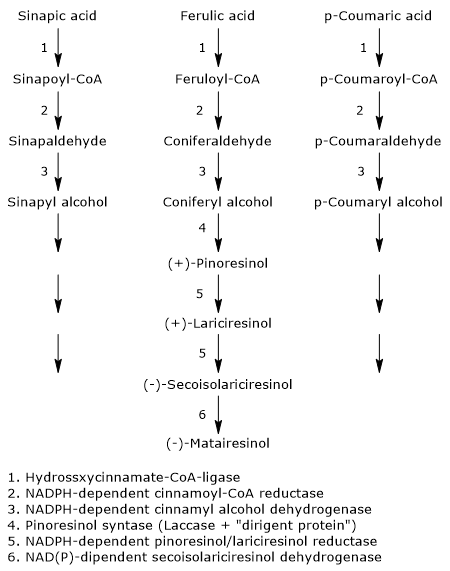
The first three reactions reduce the carboxylic group of the hydroxycinnamates to alcohol group, with formation of the corresponding alcohols, called monolignols, that is, p-coumaric alcohol, sinapyl alcohol and coniferyl alcohol. These molecules also enter the pathway of lignin biosynthesis.
- The first step, which leads to the activation of the hydroxycinnamic acids, is catalysed by hydroxycinnamate:CoA ligases, commonly called p-coumarate:CoA ligases (EC 6.2.1.12), with formation of the corresponding hydroxycinnamate-CoAs, namely, feruloil-CoA, p- coumaroyl-CoA and sinapil-CoA.
- In the second step, a NADPH-dependent cinnamoyl-CoA: oxidoreductase, also called cinnamoyl-CoA reductase (EC1.2.1.44) catalyzes the formation of the corresponding aldehydes, and the release of coenzyme A.
- In the last step, a NADPH-dependent cinnamyl alcohol dehydrogenase, also called monolignol dehydrogenase (EC 1.1.1.195), catalyzes the reduction of the aldehyde group to an alcohol group, with the formation of the aforementioned monolignols.
The next step, the dimerization of monolignols, involves the intervention of stereoselective mechanisms, or, more precisely, enantioselective mechanisms.In fact, most of the plant lignans exists as (+)- or (-)-enantiomers, that is, isomers with property of chirality, whose relative amounts can vary from species to species, but also in different organs on the same plant, depending on the type of reactions involved.
The dimerization can occur through enzymatic reactions involving laccases (EC 1.10.3.2). These enzymes catalyze the formation of radicals that, dimerizing, form a racemic mixture. However, this does not explain how the racemic mixtures found in plants are formed. The most accepted mechanism to explain the stereospecific synthesis involves the action of the laccase and of a protein able to direct the synthesis toward one or the other of the two enantiomeric forms: the dirigent protein. The reaction scheme might be: the enzyme catalyzes the synthesis of phenylpropanoid radicals that are orientated in such a way to obtain the desired stereospecific coupling by the dirigent protein.
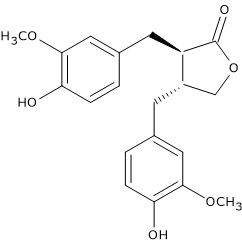
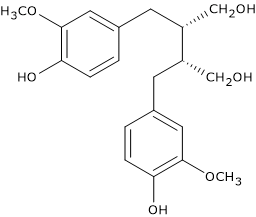
For example, pinoresinol synthase, consisting of laccase and dirigent protein, catalyzes the stereospecific synthesis of (+)-pinoresinol from two units of coniferyl alcohol. (+)-Pinoresinol, in two consecutive stereospecific reactions catalyzed by NADPH-dependent pinoresinol/lariciresinol reductase (EC 1.23.1.2), is first reduced to (+)-lariciresinol, and then to (-)-secoisolariciresinol. (-)-Secoisolariciresinol, in the reaction catalyzed by NAD(P)-dependent secoisolariciresinol dehydrogenase (EC 1.1.1.331) is oxidized to (-)-matairesinol.
Metabolism by human gut microbiota
Their importance to human health is due largely to their metabolism by gut microbiota, which is part of the larger human microbiota, and that carries out deglycosylations, para-dehydroxylations, and meta-demethylations without enantiomeric inversion.
Indeed, this metabolization leads to the formation molecules with a modest estrogen-like activity (phytoestrogens), a situation similar to that observed with some isoflavones, such as those of soybean, some coumarins, and some stilbenes. These active metabolites are the so-called “mammalian lignans or enterolignans”, such as the aglycones of enterodiol and enterolactone, formed from secoisolariciresinol and matairesinol, respectively.
Studies conducted on animals fed diets rich in lignans have shown their presence as intact molecules, in low concentrations, in serum, suggesting that they may be absorbed as such from the intestine. These molecules exhibit estrogen-independent actions, both in vivo and in vitro, such as inhibition of angiogenesis, reduction of diabetes, and suppression of tumor growth.
Note: The term “phytoestrogen” refers to molecules with estrogenic or antiandrogenic activity, at least in vitro.
Once absorbed, they enter the enterohepatic circulation, and, in the liver, may undergo phase II reactions and be sulfated or glucuronidated, and finally excreted in the urine.
Food sources
The richest dietary source is flaxseed (linseed), that contains mainly secoisolariciresinol, but also lariciresinol, pinoresinol and matairesinol in good quantity (for a total amount of more than 3.7 mg/100 g dry weight). They are also found in sesame seeds.
Another important source is whole grains.
They are also present in other foods, but in concentrations from one hundred to one thousand times lower than those of flaxseed. Examples are:
- beverages, generally more abundant in red wine, followed in descending order by black tea, soy milk and coffee;
- fruits, such as apricots, pears, peaches, strawberries;
- among vegetables, Brassicaceae, garlic, asparagus and carrots;
- lentils and beans.
Their presence in grains and, to a lesser extent in red wine and fruit, makes them, at least in individuals who follow a mediterranean diet, the main source of phytoestrogens.
References
- Andersen Ø.M., Markham K.R. Flavonoids: chemistry, biochemistry, and applications. CRC Press Taylor & Francis Group, 2006
- de la Rosa L.A., Alvarez-Parrilla E., Gonzàlez-Aguilar G.A. Fruit and vegetable phytochemicals: chemistry, nutritional value, and stability. 1th Edition. Wiley J. & Sons, Inc., Publication, 2010
- Heldt H-W. Plant biochemistry – 3th Edition. Elsevier Academic Press, 2005
- Manach C., Scalbert A., Morand C., Rémésy C., and Jime´nez L. Polyphenols: food sources and bioavailability. Am J Clin Nutr 2004;79(5):727-747. doi:10.1093/ajcn/79.5.727
- Satake H, Koyama T., Bahabadi S.E., Matsumoto E., Ono E. and Murata J. Essences in metabolic engineering of lignan biosynthesis. Metabolites 2015;5:270-290. doi:10.3390/metabo5020270
- Tsao R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010;2:1231-1246. doi:10.3390/nu2121231
- van Duynhoven J., Vaughan E.E., Jacobs D.M., Kemperman R.A., van Velzen E.J.J, Gross G., Roger L.C., Possemiers S., Smilde A.K., Doré J., Westerhuis J.A.,and Van de Wiele T. Metabolic fate of polyphenols in the human superorganism. PNAS 2011;108(suppl. 1):4531-4538. doi:10.1073/pnas.1000098107
- Wink M. Biochemistry of plant secondary metabolism – 2nd Edition. Annual plant reviews (v. 40), Wiley J. & Sons, Inc., Publication, 2010