Le proteine animali, vegetali, batteriche e virali sono grandi polimeri lineari costituiti da centinaia e talvolta anche migliaia di subunità chiamate amminoacidi.
La sequenza amminoacidica, che è unica e codificata geneticamente, è chiamata catena polipeptidica o struttura primaria. Nella catena polipeptidica, gli amminoacidi sono collegati in serie da legami covalenti chiamati legami peptidici.
La struttura primaria determina in larga misura la struttura tridimensionale, o conformazione, della proteina.
Le proteine sono una classe di molecole ampia e diversificata. Sono presenti in tutti gli organismi viventi, in tutti i compartimenti cellulari, e presentano strutture molto diverse, anche all’interno dello stesso tipo di cellula, dove si possono trovare centinaia o addirittura migliaia di proteine diverse, ciascuna delle quali svolge una funzione distinta.
La grande varietà di funzioni che sono in grado di svolgere deriva dalla capacità della catena polipeptidica di ripiegarsi in specifiche strutture tridimensionali, che conferiscono anche la capacità di legarsi a molecole diverse. Si può quindi affermare che le proteine sono il mezzo attraverso il quale si esprime l’informazione genetica.
In Sintesi: Punti Chiave
- Cosa sono: polimeri lineari composti da catene di amminoacidi uniti da legami peptidici.
- Livelli strutturali: l’organizzazione delle proteine si articola in struttura primaria, secondaria, terziaria e quaternaria.
- Motivi e Domini: le proteine globulari presentano combinazioni di strutture secondarie (motivi) e unità funzionali indipendenti (domini).
- Funzioni biologiche: agiscono come enzimi (catalizzatori), proteine strutturali (collagene), di trasporto (emoglobina) e di difesa (anticorpi).
- Determinismo: la struttura primaria, codificata geneticamente, determina la conformazione tridimensionale e la funzione specifica della proteina.
- Complessi: le proteine possono associarsi in macchine macromolecolari o complessi multienzimatici per svolgere funzioni cellulari coordinate.
Indice
Cenni storici
Grazie al fatto di essere più facili da isolare rispetto agli acidi nucleici, ai lipidi e ai polisaccaridi, lo studio delle proteine ha preceduto quello delle altre biomolecole e può essere fatto risalire ai lavori sulla composizione chimica delle albumine condotti da Jöns Jacob Berzelius, uno dei padri della chimica moderna, e Gerardus Johannes Mulder nel 1839. Per confronto, la scoperta del ruolo degli acidi nucleici nella trasmissione ed espressione dell’informazione genetica avvenne alla fine degli anni ’40 del secolo scorso, quella del loro ruolo catalitico solo negli anni ’80, mentre il ruolo dei lipidi nelle membrane biologiche venne alla luce negli anni ’60.
Il termine proteina deriva dal greco proteios, che significa “primario”, e fu suggerito da Berzelius a Mulder. Berzelius riteneva infatti che le proteine potessero essere le sostanze biologiche più importanti.
Struttura delle proteine
Al pari delle altre macromolecole biologiche, le proteine sono formate dall’unione di piccole molecole organiche, nel loro caso gli amminoacidi.
Sono stati individuati circa 20 amminoacidi differenti i quali, secondo la convenzione di Fischer, sono presenti quasi esclusivamente nella forma L. Occasionalmente, in proteine di origine batterica sono stati trovati D-amminoacidi.
Gli L-amminoacidi sono composti organici bifunzionali, in quanto presentano un gruppo carbossilico e uno amminico legati a un atomo di carbonio, detto carbonio α; per questa ragione sono anche detti L-α-amminoacidi. Ogni amminoacido è identificato da un gruppo caratteristico, detto gruppo R, anch’esso legato al carbonio α. Il gruppo R è responsabile delle proprietà chimiche dell’amminoacido, in quanto possiede una varietà di dimensioni, carica, forme e reattività proprie.
Nel corso della sintesi della proteina, il gruppo carbossilico di un amminoacido e quello amminico di un secondo sono legati covalentemente attraverso una reazione di condensazione catalizzata da specifici enzimi, ossia proteine dotate di attività catalitica. Nel corso della reazione si verifica il rilascio di una molecola d’acqua, con formazione di un legame peptidico. Si tratta di un legame rigido, planare e molto stabile in condizioni di pH fisiologico, tanto che in assenza di interventi esterni la sua vita è di circa 1.100 anni. Dal legame end-to-end di molti amminoacidi si viene a formare lo scheletro polipeptidico della proteina.
Nella descrizione della struttura che assume nello spazio la catena polipeptidica è di aiuto distinguere vari livelli di organizzazione, ossia la struttura primaria, secondaria, le strutture supersecondarie, i domini, la struttura terziaria e quella quaternaria.
Nota: la convenzione di Fischer non è più utilizzata, se non per carboidrati e amminoacidi, sostituita dal sistema RS, il quale consente di assegnare in maniera più accurata il nome a molecole dotate di un centro di chiralità.
Struttura primaria
La prima proteina di cui è stata determinata la struttura primaria fu l’insulina, grazie al lavoro di Frederick Sanger.
La struttura primaria è la sequenza amminoacidica di una proteina, il suo livello di organizzazione più basso, e, come detto, è unica, geneticamente determinata, e ad essa si deve la struttura tridimensionale e la funzione della proteina. La struttura primaria può essere formata da 40 fino a oltre 4.000 residui amminoacidici.
La catena polipeptidica è dotata di polarità in quanto le due estremità sono differenti: una presenta un gruppo amminico libero, detto gruppo ammino-terminale, l’altra un gruppo carbossilico libero, detto gruppo carbossi-terminale. Le due estremità della catena polipeptidica sono definite anche estremità N-terminale e C-terminale, per distinguere i due gruppi da quelli presenti nelle catene laterali degli amminoacidi interni alla catena. Per convenzione, l’estremità amminica libera è considerata l’inizio della catena, e si scrive a sinistra.
La struttura primaria è interessante anche perché, confrontando quella di una stessa proteina presente in specie differenti, si possono individuare le variazioni che il gene corrispondente ha subito, che sono un indice di quanto le specie si siano separate nel corso dell’evoluzione.
I termini dipeptide, tripeptide, oligopeptide e polipeptide sono utilizzati per indicare catene di lunghezza diversa, rispettivamente composte da 2, 3, meno di 50, e più di 50 residui.
Struttura secondaria
La scoperta della struttura secondaria delle proteine si deve al lavoro di Linus Pauling e Robert Corey, i quali proposero l’esistenza di due strutture dette α-elica e foglietto β o struttura β a foglietto pieghettato.
La struttura secondaria deriva dalla formazione di legami a idrogeno tra parti contigue della catena polipeptidica dotate di particolari sequenze amminoacidiche. Dunque, descrive la disposizione nello spazio di residui amminoacidici adiacenti lungo la struttura primaria.
Oltre alle strutture sopra citate ne sono state individuate altre quali i β turn, i γ turn e gli Ω loop, tutte appartenenti al gruppo definito reverse turns. Queste strutture sono spesso presenti nei punti dove la catena polipeptidica inverte la propria direzione, e in genere sono disposte sulla superficie della molecola.
Circa il 32-38% degli amminoacidi delle proteine globulari si ritrova in una conformazione ad α-elica.
Le strutture successive a quella secondaria sono presenti solo nelle proteine globulari.
Strutture supersecondarie o motivi
Le strutture supersecondarie sono combinazioni di strutture secondarie che formano una regione della molecola con una topologia e struttura tridimensionale caratteristica. Sono connesse tra di loro da regioni ad ansa prive di una struttura definita.
Tra i motivi più comuni si ritrovano l’α-α corner, il β-β corner, il β-β hairpin, il β-α-β-motif, quest’ultimo spesso presente nelle proteine che legano l’RNA o il DNA, e il 3β-corner.
Domini
I domini sono regioni globulari che risultano dalla combinazione di motivi che si ripiegano in modo indipendente dal resto della catena polipeptidica a dare una struttura stabile.
Sono formati da 40−400 amminoacidi, tranne i domini motori e chinasici che presentano un numero molto maggiore di amminoacidi.
I domini sono stati classificati in tre gruppi principali, individuati in base alle strutture secondarie e motivi presenti:
- domini α;
- domini β;
- domini α/β.
Sono state identificate oltre 1.000 famiglie di domini, e i membri di ciascuna famiglia sono detti omologhi.
Molto spesso un dominio è dotato di una specifica funzione, quindi è un’unità funzionale della proteina in cui è contenuto.
Le proteine possono essere costituite da un singolo dominio, quelle più piccole, o da più domini. Ad esempio, la chimotripsina (EC 3.4.21.1), un enzima coinvolto nella digestione delle proteine, è composta da un dominio, mentre la papaina (EC 3.4.22.2) da due domini.
Struttura terziaria
La struttura terziaria, anche detta struttura nativa, è la struttura tridimensionale della proteina ed è la sua forma biologicamente attiva.
La prima proteina di cui è stata determinata tale struttura è stata la mioglobina, grazie al lavoro di John Kendrew.
La catena polipeptidica non è una struttura rigida ed è capace di ripiegarsi spontaneamente a dare una struttura tridimensionale, che in gran parte è determinata proprio dalla sua sequenza amminoacidica. Il ripiegamento della struttura primaria permette la transizione dal mondo unidimensionale della struttura primaria a quello tridimensionale della proteina.
In questo tipo di struttura, il ripiegamento della catena polipeptidica fa sì che residui amminoacidici lontani si ritrovino vicini, dunque riguarda la disposizione nello spazio di amminoacidi lontani tra di loro nella struttura primaria.
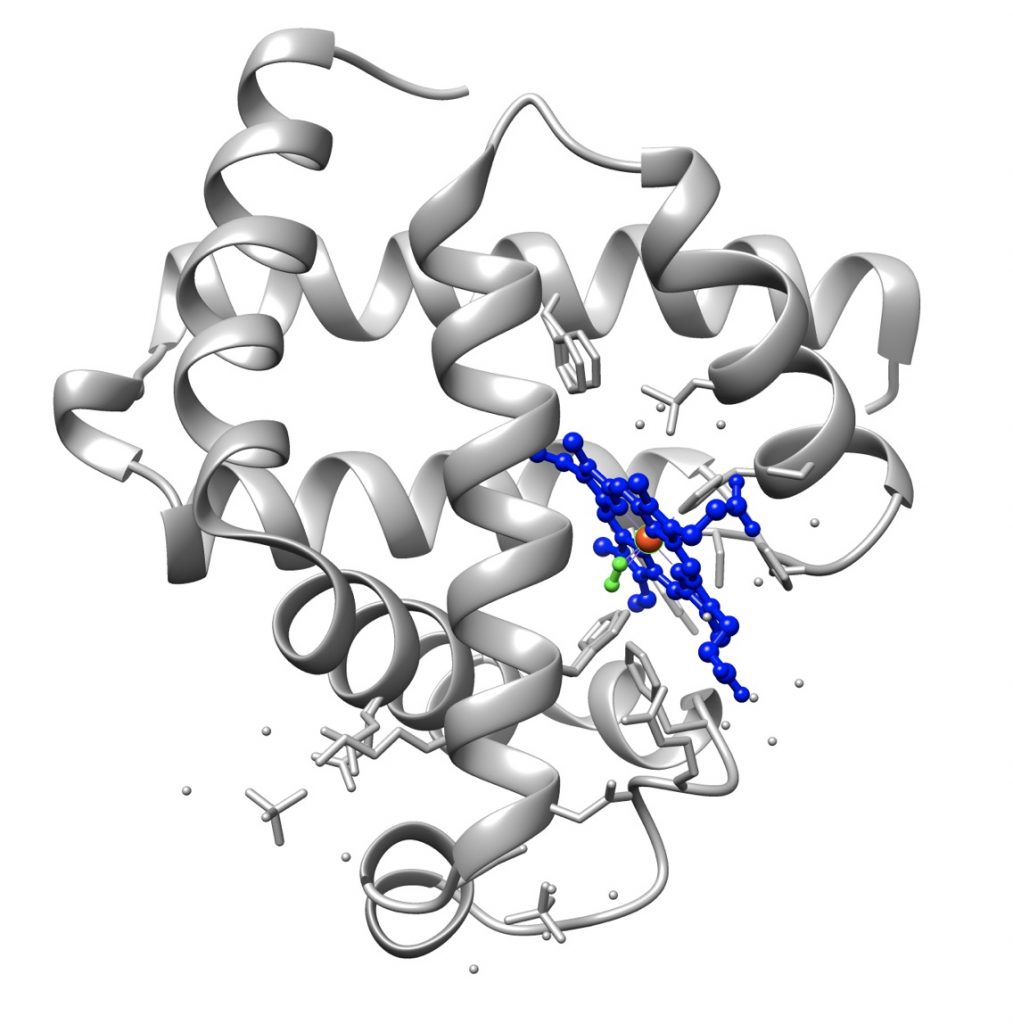
La struttura terziaria, specie delle proteine formate da più di 200 residui amminoacidici, è formata da diversi domini uniti da brevi segmenti polipeptidici, ed è spesso stabilizzata da legami disolfuro tra residui di cisteina, che si stabiliscono dopo che la molecola ha raggiunto la sua struttura nativa.
Non tutte le proteine globulari sono dotate di struttura terziaria. Un esempio sono le caseine del latte, la cui catena polipeptidica assume una conformazione tridimensionale disordinata, anche detta random coiled structure. La struttura tridimensionale disordinata fa sì che queste proteine siano molto suscettibili all’attacco delle proteasi intestinali, e quindi alla liberazione degli amminoacidi costituenti, il che le rende estremamente idonee alla loro funzione nutrizionale. Un altro esempio di proteina random coiled è l’elastina, una delle più abbondanti proteine dell’organismo.
Struttura quaternaria
Questo ulteriore livello di organizzazione strutturale descrive come due o più catene polipeptidiche si associano a dare una singola struttura proteica. Dunque si riferisce alla disposizione spaziale delle singole catene e alla natura delle forze che le legano, quali:
- l’effetto idrofobo, che è anche il principale responsabile del ripiegamento delle proteine;
- i legami idrogeno;
- le interazioni di van der Waals;
- le interazioni ioniche;
- i legami covalenti trasversali.
La struttura risultante è definita oligomero o proteina oligomerica, e le subunità polipeptidiche costituenti, che possono essere uguali o diverse, sono chiamate monomeri o semplicemente subunità.
La maggior parte delle proteine intracellulari sono oligomeri, a differenza della maggior parte di quelle extracellulari. Un esempio di proteina con struttura quaternaria è l’emoglobina.
Questo livello di struttura è ovviamente assente nelle proteine monomeriche formate da una sola catena polipeptidica.
Le proteine sono anche in grado di interagire tra di loro a formare strutture, dette macchine macromolecolari, nelle quali, agendo in modo sinergico, riescono a svolgere funzioni che da sole non sarebbero capaci di portare a termine. Esempi sono i complessi multienzimatici, come il complesso della piruvato deidrogenasi.
Funzioni delle proteine
Le proteine svolgono una gamma straordinariamente ampia di funzioni biologiche, che riflettono sia la versatilità strutturale della catena polipeptidica sia la diversità chimica delle catene laterali degli amminoacidi. Le loro funzioni derivano dalla capacità delle proteine di ripiegarsi in specifiche conformazioni tridimensionali e di interagire selettivamente con altre molecole, tra cui piccoli metaboliti, macromolecole e ioni.
Da un punto di vista funzionale, le proteine partecipano praticamente a tutti i processi biologici. Agiscono come:
- enzimi, catalizzando reazioni biochimiche essenziali per il metabolismo;
- proteine di trasporto, consentendo il movimento di sostanze come ossigeno, lipidi e ioni metallici all’interno dell’organismo;
- componenti strutturali, fornendo supporto meccanico a cellule e tessuti;
- molecole regolatrici, controllando processi come l’espressione genica, la trasduzione del segnale, la crescita e la differenziazione.
Classificazione dei ruoli biologici
Data questa diversità funzionale, l’approccio più efficace per comprendere le proteine è quello di classificarle in base agli specifici ruoli biologici che svolgono all’interno della cellula.
| Categoria funzionale | Esempi tipici e dettagli | Ruolo biologico e meccanismo principale |
|---|---|---|
| Enzimi | DNA polimerasi, α-amilasi | Catalizzatori biologici che accelerano le reazioni biochimiche vitali per il metabolismo. |
| Proteine di trasporto | Emoglobina, GLUT4, transferrina | Consentono il movimento selettivo di sostanze (ossigeno, glucosio, ioni) nell’organismo. |
| Proteine strutturali | Collagene, cheratina, elastina | Forniscono supporto meccanico, elasticità e resistenza a cellule, tessuti e organi. |
| Proteine di difesa | Immunoglobuline (IgG), sistema del complemento | Proteggono l’organismo da agenti patogeni e supportano la risposta immunitaria. |
| Proteine contrattili e motorie | Actina, miosina, chinesina | Responsabili della generazione del movimento, della contrazione muscolare e del trasporto cellulare. |
| Proteine di regolazione | Insulina, fattori di trascrizione | Controllano e coordinano i processi biologici, l’espressione genica e la trasduzione del segnale. |
| Fonte di energia | Amminoacidi da catabolismo proteico | Forniscono substrati alternativi quando gli altri macronutrienti sono insufficienti, ad esempio durante il digiuno. |
Questa classificazione funzionale fornisce un quadro utile per comprendere come la struttura delle proteine sia strettamente correlata all’attività biologica. Di fatto, l’inquadramento basato sui ruoli biochimici rappresenta uno dei pilastri fondamentali nella classificazione delle proteine.
Bibliografia
- Alberts B., Johnson A., Lewis J., Morgan D., Raff M., Roberts K., Walter P. Molecular biology of the cell. 7th Edition. Garland Science, Taylor & Francis Group, 2022.
- Berg J.M., Tymoczko J.L., Gatto J.G., Stryer L. Biochemistry. 9th Edition. W.H. Freeman and Company, 2019.
- Branden C., Tooze J. Introduction to protein structure. 2nd Edition. Garland Science, 1999. doi:10.1201/9781136969898
- Garrett R.H., Grisham C.M. Biochemistry. 7th Edition. Cengage Learning, 2023.
- Heilman D., Woski S., Voet D., Voet J.G., Pratt C.W. Fundamentals of biochemistry: life at the molecular level. 6th Edition. Wiley, 2023.
- Kessel A., Ben-Tal N. Introduction to proteins: structure, function, and motion. CRC Press, 2011.
- Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 8th Edition. W.H. Freeman and Company, 2021.
- Petsko G.A., Ringe D. Protein structure and function. Oxford University Press, 2008.
- Rodwell V.W., Bender D.A., Botham K.M., Kennelly P.J., Weil P.A. Harper’s illustrated biochemistry. 31st Edition. McGraw-Hill, 2018.
- Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 4th Edition. St. Louis: Elsevier, 2018.
Domande Frequenti
Cosa sono le proteine e da che cosa sono composte?
Le proteine sono grandi polimeri lineari costituiti da catene di centinaia o migliaia di subunità chiamate amminoacidi. Questi ultimi sono legati in serie da legami covalenti molto stabili, detti legami peptidici, che formano lo scheletro della macromolecola.
Perché la struttura primaria di una proteina è così importante?
La struttura primaria è la sequenza amminoacidica unica e codificata geneticamente. Questo livello determina il modo in cui la catena polipeptidica si ripiega nello spazio, stabilendo la conformazione tridimensionale finale e la specifica funzione biologica.
Qual è la differenza tra motivi e domini proteici?
I motivi sono semplici combinazioni di strutture secondarie vicine. I domini sono invece regioni globulari più grandi e complesse, formate dall'unione di più motivi, capaci di ripiegarsi in modo indipendente e dotate di una specifica funzione.
Quali sono le principali funzioni biologiche delle proteine?
Le proteine svolgono ruoli straordinari: agiscono come enzimi per accelerare le reazioni, come trasportatori di sostanze (emoglobina), come componenti strutturali (collagene) e come molecole di difesa (anticorpi) o regolatrici per il controllo dell'espressione genica.