When excess calories are consumed from carbohydrates or proteins, such surplus is used to synthesize fatty acids and then triacylglycerols, while it doesn’t occur if the excess come from fats.
Contents
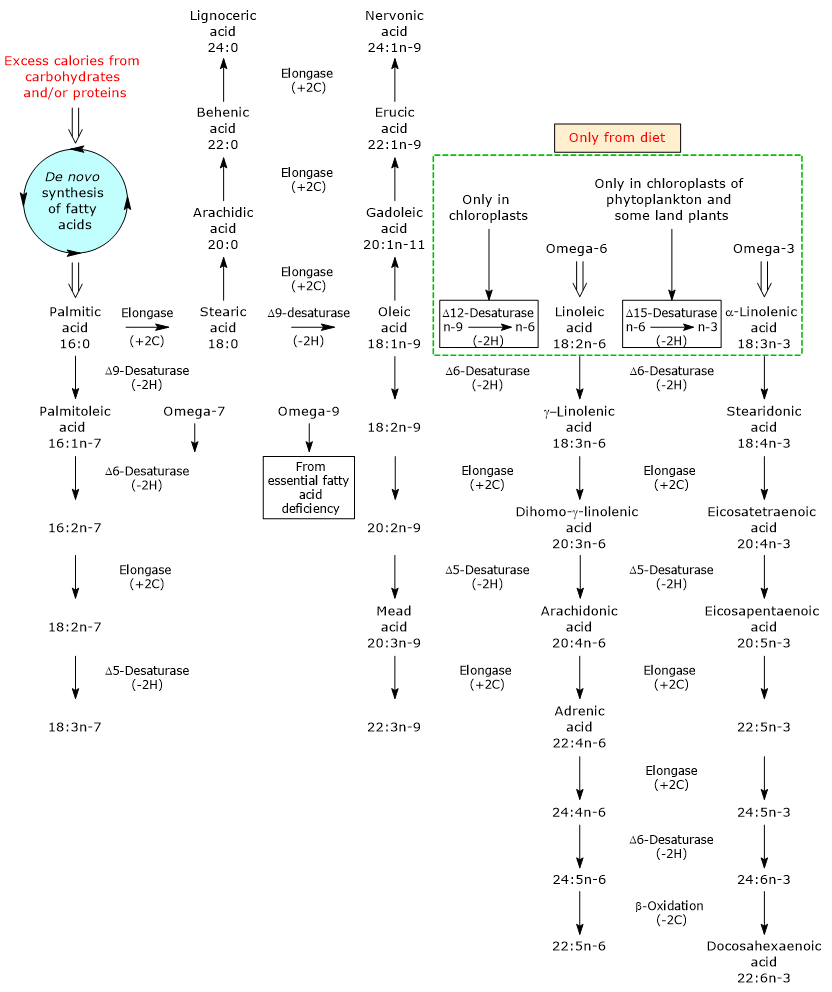
De novo fatty acid synthesis in plants and animals
De novo fatty acid synthesis is largely similar among plants and animals. It occurs in chloroplasts of photosynthetic cells of higher plants, and in cytosol of animal cells by the concerted action of two enzymes: acetyl-CoA carboxylase (EC 6.4.1.2) and fatty acid synthase (EC 2.3.1.85).
Fatty acid synthase catalyzes a repeating four-step sequence by which the fatty acyl chain is extended by two carbons, at the carboxyl end, every each passage through the cycle; this four-step process is the same in all organisms.
In animals, the primary site for lipid metabolism is liver, not the adipose tissue. However, adipose tissue is a major organ system in which fatty acid synthesis occurs, though in humans it is less active than in many other animal species.
Although myristic acid, lauric acid and a trace of stearic acid may also be produced, in animals and plants the main product of these reactions is palmitic acid.
It should be noted that in certain plants, such as palm and coconut, chain termination occurs earlier than palmitic acid release: up to 90 percent of the fatty acids produced and then present in the oils of these plants are between 8 (caprylic acid) and 14 (myristic acid) carbons long (palmitic acid: 16 carbon atoms).
Synthesis of long chain saturated and unsaturated fatty acids
Palmitic acid is the commonest saturated fatty acid in plant and animal lipids. Generally it is not present in very large proportions because it may be undergo into several metabolic pathways.
In fact:
- it is the precursor of stearic acid;
- it may be desaturated to palmitoleic acid, the precursor of all fatty acids of omega-7 or n-7 family, in a reaction catalyzed by Δ9-desaturase (EC 1.14.19.1), an ubiquitous enzyme in both plant and animal kingdoms and the most active lipid enzyme in mammalian tissues, the same enzyme that catalyzes the desaturation of stearic acid to oleic acid. Note: Δ9-desaturase inserts double bounds in the 9-10 position of the fatty acid carbon chain, position numbered from the carboxyl end of the molecule, and:
if the substrate is palmitic acid, the double bond is inserted between n-7 and n-8 position of the chain, so producing palmitoleic acid, the founder of omega-7 series;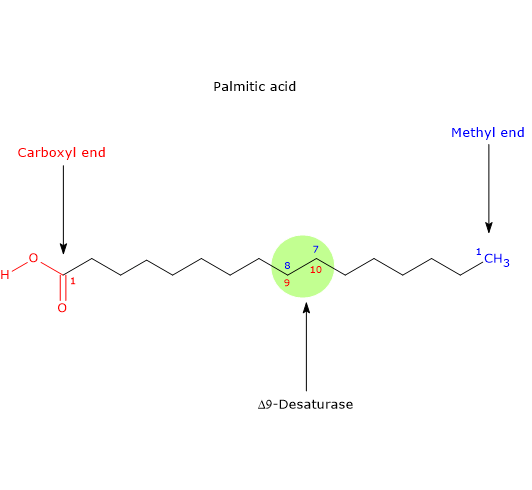
if the substrate is stearic acid, the double bond is inserted between n-9 and n-10 position of the chain and oleic acid will be produced.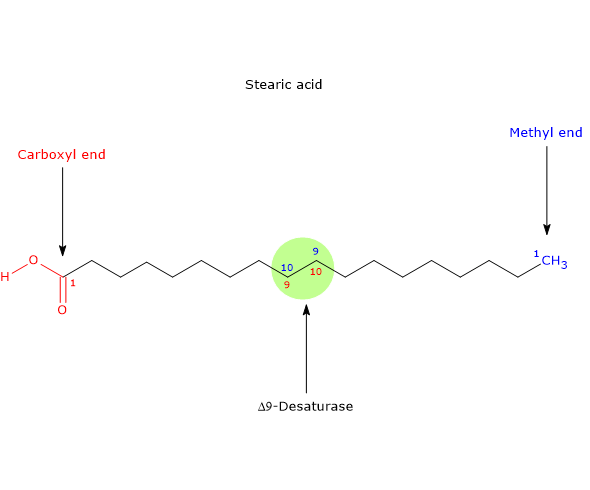
- It may be esterified into complex lipids.
Of course, in plants and animals there are fatty acids longer and/or more unsaturated than these just seen thanks to modification systems that catalyze reactions of fatty acid synthesis that are organism- tissue- and cell- specific.
For example, stearic acid may be:
- elongated to arachidic acid, behenic acid and lignoceric acid, all saturated fatty acids, in reactions catalyzed by elongases. Again, chain elongation occurs, both in mitochondria and in the smooth endoplasmic reticulum, by the addition of two carbon atom units at a time at the carboxylic end of the fatty acid through the action of fatty acid elongation systems (particularly long and very long saturated fatty acids, from 18 to 24 carbon atoms, are synthesized only on cytosolic face of the smooth endoplasmic reticulum);
- desaturated, as seen, to oleic acid, an omega-9 or n-9 fatty acid, in a reaction catalyzed by Δ9-desaturase.
Several researchers have postulated that the reason for which stearic acid does not concur to hypercholesterolemia is its rapid conversion to oleic acid.
Oleic acid is the start point for the synthesis of many other unsaturated fatty acids by reactions of elongation and/or desaturation.
In fact:
- gadoleic acid, erucic acid and nervonic acid, all monounsaturated fatty acids. Saturated fatty acids, and unsaturated fatty acids of the omega-9 series, usually oleic acid (but also palmitoleic acid and other omega-7 fatty acids) are the only fatty acids produced de novo in mammal systems.
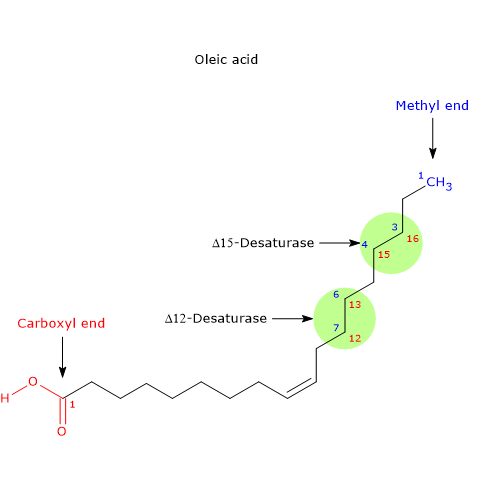
- Thanks to the consecutive action of the enzymes Δ12-desaturase (1.14.19.6) and Δ15-desaturase (EC 1.14.19.25), that insert a double bond respectively in the 12-13 and 15-16 position of the carbon chain of the fatty acid, oleic acid is converted first to linoleic acid, founder of all the omega-6 polyunsaturated fatty acids, and then to alpha-linolenic acid, founder of all the omega-3 polyunsaturated fatty acids (omega-3 and omega-6 PUFA that will be produced from these precursors through repeating reactions of elongation and desaturation).
- In the case of essential fatty acid (EFA) deficiency, oleic acid may be desaturated and elongated to omega-9 polyunsaturated fatty acids, with accumulation especially of Mead acid.
Omega-3 and omega-6 PUFA synthesis
Animal tissues can desaturate fatty acids in the 9-10 position of the chain, thanks to the presence of Δ9 desaturase; as previously seen, if the substrate of the reaction is palmitic acid, the double bond will appear between n-7 and n-8 position, with stearic acid between n-9 and n-10 position, so leading to formation respectively of palmitoleic acid and oleic acid.
Animals lack Δ12- and Δ15-desaturases, enzymes able to desaturate carbon carbon bonds beyond the 9-10 position of the chain. For these reason, they can’t produce de novo omega-3 and omega-6 PUFA (which have double bonds also beyond the 9-10 position), that are so essential fatty acids.
Δ12- and Δ15-desaturases are present in plants; though many land plants lack Δ15-desaturase, also called omega-3 desaturase, planktons and aquatic plants in colder water possess it and produce abundant amounts of the omega-3 fatty acids.
References
- Akoh C.C. and Min D.B. “Food lipids: chemistry, nutrition, and biotechnology”. CRC Press Taylor & Francis Group, 2008 3th ed. 2008
- Bender D.A. “Benders’ dictionary of nutrition and food technology”. 2006, 8th Edition. Woodhead Publishing. Oxford
- Burr G.O. and Burr M.M. A new deficiency disease produced by the rigid exclusion of fat from the diet. Nutr Rev 1973;31(8):148-149. doi:10.1111/j.1753-4887.1973.tb06008.x
- Chow Ching K. “Fatty acids in foods and their health implication”. 3rd Edition. CRC Press Taylor & Francis Group, 2008
- Rosenthal M.D., Glew R.H. Mediacal biochemistry. Human metabolism in health and disease. John Wiley & Sons, Inc. 2009
- Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 3rd Edition. Elsevier health sciences, 2012