

Glucogenic amino acids are defined as amino acids whose carbon skeletons can be fully or partially catabolized into precursors for gluconeogenesis, hence the name “glucogenic”.[1]
Unlike glycogen or triglycerides, the body does not store excess amino acids.[2]
When cellular protein and energy needs are met, surplus amino acids are catabolized into precursors for the synthesis of fatty acids, ketone bodies, or glucose.[3]
Glucogenic and ketogenic amino acids are classified based on the metabolic fate of their carbon skeletons. However, this classification is not clear-cut because, among the twenty standard amino acids found in proteins, five are both glucogenic and ketogenic.[4]
Glucose synthesis is possible because their carbon skeletons yield pyruvate, oxaloacetate, α-ketoglutarate, succinyl-CoA, or fumarate.[5]
While pyruvate and oxaloacetate directly enter gluconeogenesis, the other intermediates provide a net gain of carbon units within the citric acid cycle, eventually forming oxaloacetate.[6]
Consequently, during prolonged fasting or carbohydrate restriction, glucogenic amino acids play a crucial role, serving as a major source of precursors for gluconeogenesis, second only to glycerol derived from adipose tissue lipolysis.[7]
Summary: Key Points
- Definition: amino acids whose carbon skeletons are catabolized into precursors for glucose synthesis (gluconeogenesis).
- Classification: out of 20 standard amino acids, 13 are exclusively glucogenic and 5 are mixed (both glucogenic and ketogenic).
- Metabolic entry: converted into pyruvate or citric acid cycle intermediates, leading to a net gain of carbon units for gluconeogenesis.
- Acetyl-CoA block: Acetyl-CoA cannot serve as a glucose precursor due to the irreversible nature of the pyruvate dehydrogenase reaction in animals.
- Prolonged fasting: Alongside glycerol, they are the primary source for maintaining blood glucose once glycogen stores are depleted.
- Alanine transport: Alanine serves as the predominant hepatic gluconeogenic substrate, carried from muscles via the glucose-alanine cycle.
Contents
- What are glucogenic amino acids?
- Biochemical basis
- Glucogenic amino acids during prolonged fasting
- References
What are glucogenic amino acids?
Of the twenty amino acids that make up proteins, thirteen are exclusively glucogenic, meaning that the catabolism of their carbon skeletons yields only glucose precursors.[4]
| Amino acid | Ketogenic | Glucogenic | |
|---|---|---|---|
| Acetyl-CoA | Acetoacetyl-CoA | ||
| Alanine | ✔ Pyruvate | ||
| Arginine | ✔ α-Ketoglutarate | ||
| Asparagine | ✔ Fumarate | ||
| Aspartate | ✔ Oxaloacetate | ||
| Cysteine | ✔ Pyruvate | ||
| Phenylalanine * | ✔ | ✔ Fumarate | |
| Glycine | ✔ Pyruvate | ||
| Glutamate | ✔ α-Ketoglutarate | ||
| Glutamine | ✔ α-Ketoglutarate | ||
| Isoleucine * | ✔ | ✔ Succinyl-CoA | |
| Histidine | ✔ α-Ketoglutarate | ||
| Methionine | ✔ Succinyl-CoA | ||
| Proline | ✔ α-Ketoglutarate | ||
| Serine | ✔ Pyruvate | ||
| Tyrosine * | ✔ | ✔ Fumarate | |
| Threonine * | ✔ | ✔ Succinyl-CoA | |
| Tryptophan * | ✔ | ✔ | ✔ Pyruvate |
| Valine | ✔ Succinyl-CoA | ||
| * Amino acids that are both glucogenic and ketogenic. | |||
Five amino acids, namely isoleucine, phenylalanine, threonine, tryptophan, and tyrosine, are both glucogenic and ketogenic, as a portion of their carbon skeleton is catabolized into glucogenic precursors, while another portion is converted into acetyl-CoA and/or acetoacetyl-CoA.[8]
Before amino acid carbon skeletons can be utilized, their amino groups must first be removed. Alanine and glutamate (or glutamine), are two of the primary molecules responsible for transporting amino groups from extrahepatic tissues to the liver, are particularly important glucogenic amino acids in mammals. Alanine is the primary gluconeogenic substrate for the liver, reaching it from muscle and other peripheral tissues via the glucose-alanine cycle.[9]
Biochemical basis
As with ketogenic amino acids, analyzing the stoichiometry of the citric acid cycle helps explain why the carbon skeletons of glucogenic amino acids act as precursors for glucose synthesis. The key factor lies in the specific point at which these carbons enter the cycle.[6]

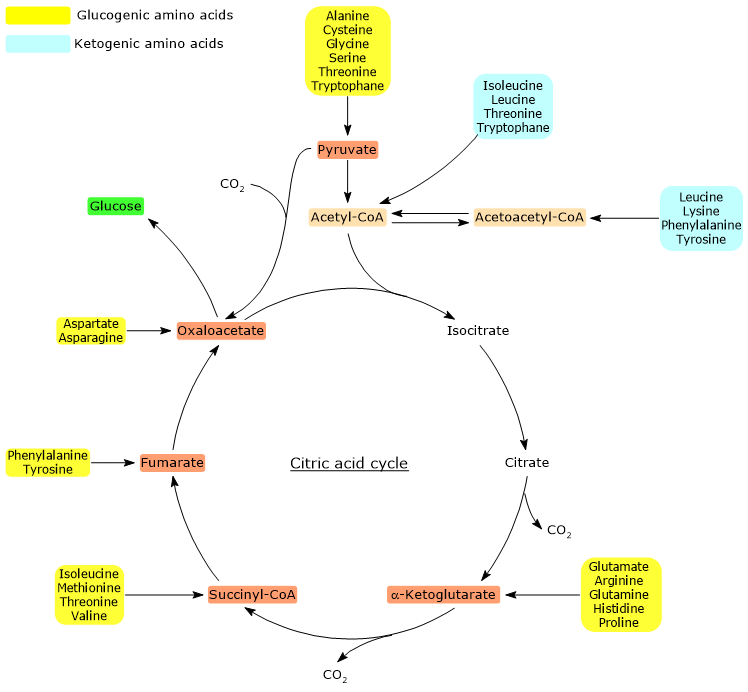
When carbon atoms derived from amino acids enter the cycle as α-ketoglutarate, succinyl-CoA, or fumarate, they contribute to a net gain of carbon units. Except for α-ketoglutarate, these intermediates enter the cycle downstream of the two oxidative decarboxylation steps catalyzed by isocitrate dehydrogenase (EC 1.1.1.42) and the α-ketoglutarate dehydrogenase multienzyme complex.[5]
This results in a net gain of one carbon unit when entry occurs at α-ketoglutarate, or two when it occurs at succinyl-CoA or fumarate. These carbon atoms can then feed into gluconeogenesis via oxaloacetate.[4]
Furthermore, because the reaction catalyzed by the pyruvate dehydrogenase complex, the oxidative decarboxylation of pyruvate to acetyl-CoA, is irreversible, and since animals lack a pathway to convert acetyl-CoA back to pyruvate, acetyl-CoA cannot serve as a glucogenic substrate.[6]
Glucogenic amino acids during prolonged fasting
Glucogenic amino acids play a crucial role during prolonged fasting and in diets with severe carbohydrate restriction. Under these conditions, they are among the primary precursors for glucose synthesis.[6] The utilization of their carbon skeletons, along with glycerol and propionate, a short-chain fatty acid, helps maintain glycemic homeostasis via gluconeogenesis when liver glycogen stores are depleted.[7]
However, even under physiological conditions, such as during cellular protein turnover or after a protein-rich meal, amino acids exceeding the requirements for protein synthesis are catabolized. Depending on the metabolic state and the specific amino acid, they may be used for energy production or anabolic processes, including glucose synthesis. The glucose produced can subsequently be utilized for glycogen synthesis, converted into ketone bodies or fatty acids.[2]
References
- ^ D’Andrea G. Classifying amino acids as gluco(glyco)genic, ketogenic, or both. Biochem Educ 2000;28(1):27-28. doi:10.1016/s0307-4412(98)00271-4
- ^ a b Brosnan J.T. Interorgan amino acid transport and its regulation. J Nutr 2003;133(6 Suppl 1):2068S-2072S. doi:10.1093/jn/133.6.2068S
- ^ Litwack G. Human biochemistry. 2nd Edition. Academic Pr, 2021.
- ^ a b c Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 8th Edition. W.H. Freeman and Company, 2021.
- ^ a b Moran L.A., Horton H.R., Scrimgeour K.G., Perry M.D. Principles of Biochemistry. 5th Edition. Pearson, 2012.
- ^ a b c d Heilman D., Woski S., Voet D., Voet J.G., Pratt C.W. Fundamentals of biochemistry: life at the molecular level. 6th Edition. Wiley, 2023.
- ^ a b Kuriyama H., Shimomura I., Kishida K., Kondo H., Furuyama N., Nishizawa H., Maeda N., Matsuda M., Nagaretani H., Kihara S., Nakamura T., Tochino Y., Funahashi T., Matsuzawa Y. Coordinated regulation of fat-specific and liver-specific glycerol channels, aquaporin adipose and aquaporin 9. Diabetes 2002;51(10):2915-2921. 10.2337/diabetes.51.10.2915
- ^ Rosenthal M.D., Glew R.H. Medical Biochemistry – Human Metabolism in Health and Disease. John Wiley J. & Sons, Inc., 2009.
- ^ Felig P., Pozefsk T., Marlis E., Cahill G.F. Alanine: key role in gluconeogenesis. Science 1970;167(3920):1003-1004. doi:10.1126/science.167.3920.1003